- Home
- About Journals
-
Information for Authors/ReviewersEditorial Policies
Publication Fee
Publication Cycle - Process Flowchart
Online Manuscript Submission and Tracking System
Publishing Ethics and Rectitude
Authorship
Author Benefits
Reviewer Guidelines
Guest Editor Guidelines
Peer Review Workflow
Quick Track Option
Copyediting Services
Bentham Open Membership
Bentham Open Advisory Board
Archiving Policies
Fabricating and Stating False Information
Post Publication Discussions and Corrections
Editorial Management
Advertise With Us
Funding Agencies
Rate List
Kudos
General FAQs
Special Fee Waivers and Discounts
- Contact
- Help
- About Us
- Search




The Open Virology Journal
(Discontinued)
ISSN: 1874-3579 ― Volume 15, 2021
Molecular and Serological Evidence for the Presence of Novel Phleboviruses in Sandflies from Northern Algeria
Grégory Moureau1, Laurence Bichaud1, Nicolas Salez1, Laetitia Ninove1, Boussad Hamrioui2, Smail Belazzoug3, Xavier de Lamballerie1, Arezki Izri4, Rémi N Charrel*, 1
Abstract
During summer 2007, a total of 785 phlebotomine flies were trapped in northern Algeria, identified morphologically, organised as monospecific pools and tested for the presence of phlebovirus RNA using degenerate primers. Three pools were positive, and the corresponding PCR products were cloned and sequenced. Viral sequences corresponding to two phleboviruses distinct from each other were detected in sandflies circulating in two close locations (140 km apart) in Northern Algeria. The 3 sequences were aligned with homologous polymerase sequences retrieved from the Genbank database, in order to examine their phylogenetic relationships. One viral sequence (from Phlebotomus papatasi) was closely related to but distinct from a sequence obtained from Phlebotomus ariasi sandflies trapped in Algeria in 2006. The two other viral sequences (from Phlebotomus longicuspis) were genetically distantly related to sequences corresponding to virus members of the Sandfly fever Naples virus species and although falling within the same group, this clearly represents a second distinct novel lineage. These results are indicative of a high genetic heterogeneity within sandflies trapped in a relatively small geographic area. Seroprevalence studies conducted on sera from populations living in the same areas indicated that humans can be infected by these viruses.
Article Information
Identifiers and Pagination:
Year: 2010Volume: 4
First Page: 15
Last Page: 21
Publisher Id: TOVJ-4-15
DOI: 10.2174/1874357901004010015
Article History:
Received Date: 23/11/2009Revision Received Date: 30/11/2009
Acceptance Date: 31/12/2009
Electronic publication date: 22/04/2010
Collection year: 2010
open-access license: This is an open access article licensed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted, non-commercial use, distribution and reproduction in any medium, provided the work is properly cited.
* Address correspondence to this author at the Unite des Virus Emergents, UMR190 "Emergence des Pathologies Virales, Faculte de Medecine, 27, blvd Jean Moulin, 13005 Marseille, France; Tel: +33 (0)4 91 32 44 20; Fax: +33 (0)4 91 32 44 21; E-mail: remi.charrel@univmed.fr
| Open Peer Review Details | |||
|---|---|---|---|
| Manuscript submitted on 23-11-2009 |
Original Manuscript | Molecular and Serological Evidence for the Presence of Novel Phleboviruses in Sandflies from Northern Algeria | |
INTRODUCTION
Recent field and experimental studies have provided increasing evidence that the number of known viruses in the genus Phlebovirus (Bunyaviridae family) may be largely underestimated as suggested by phylogram shape-based analyses [1Pybus OG, Rambaut A, Holmes EC, Harvey PH. New inferences from tree shape: numbers of missing taxa and population growth rates Syst Biol 2002; 51: 881-.-3Izri A, Temmam S, Moureau G, Hamrioui B, de Lamballerie X, Charrel RN. Sandfly fever Sicilian virus, Algeria Emerg Infect Dis 2008; 14: 795-.]. Until recently, sandflies distributed in the old world were known as vectors for few phleboviruses including two known species (Sandfly fever Naples virus, and Salehebad virus), and tentative species such as sandfly fever Sicilian virus and Corfou virus.
During the last decade, Toscana virus, a member of the species Sandfly fever Naples virus (SFNV), was increasingly investigated in patients presenting with febrile neurological manifestations (meningitis, encephalitis) during the summer in Mediterranean countries. The focus on Toscana virus occurred in the mid 1990's with several studies from central Italy where this virus was shown to be the most frequent cause of meningitis in central Italy during summertime [4Braito A, Ciufolini MG, Pippi L, et al. Phlebotomus-transmitted toscana virus infections of the central nervous system: a seven-year experience in Tuscany Scand J Infect Dis 1998; 30: 505-8.-6Valassina M, Meacci F, Valensin PE, Cusi MG. Detection of neurotropic viruses circulating in Tuscany: the incisive role of Toscana virus J Med Virol 2000; 60: 86-90.]. The second event which highlighted Toscana virus was the systematic surveillance initiated in many South-European countries to track West Nile virus after it invaded the United States in 1999 and caused outbreaks in Europe and North Africa [7Briese T, Jia XY, Huang C, Grady LJ, Lipkin WI. Identification of a Kunjin/West Nile-like flavivirus in brains of patients with New York encephalitis Lancet 1999; 354: 1261-2., 8Granwehr BP, Lillibridge KM, Higgs S, et al. West Nile virus: where are we now? Lancet Infect Dis 2004; 4: 547-6.]. Surveillance for West Nile virus infections demonstrated that Toscana virus, whose prevalence was also assessed by reference laboratories, (i) was much more prevalent than West Nile virus, (ii) much more prevalent than initially believed, (iii) and circulated and caused human infections in Spain, France, and Portugal, and probably many other countries where sandflies are distributed [9Mendoza-Montero J, Gamez-Rueda MI, Navarro-Mari JM, de la Rosa-Fraile M, Oyonarte-Gomez S. Infections due to sandfly fever virus serotype Toscana in Spain Clin Infect Dis 1998; 27: 434-6.-15Santos L, Simoes J, Costa R, Martins S, Lecour H. Toscana virus meningitis in Portugal, 2002-2005 Euro Surveill 2007; 12: E3-4.].
Sandfly fever Sicilian virus (SFSV) is a distinct arthropod-borne phlebovirus transmitted by sandflies, specifically by Phlebotomus papatasi [16Karabatsos N. International Catalogue of Arthropod-borne viruses In: American Society for Tropical Medicine and Hygiene. 3rd. San Antonio, Texas 1985; (Suppl 1): pp. 137-52.]. It was discovered in Italy (Palerma, Sicilia) where it affected the troops of the Allied Army Forces after the Sicily landings in 1943 during World War II. Clinical presentation is a febrile illness during the warm season; in contrast with Toscana virus infection, SFSV has not been associated with neurological manifestations. Recently, molecular evidence for the presence of a phlebovirus closely related to Sandfly fever Sicilian virus (SFSV) in Algeria was reported [3Izri A, Temmam S, Moureau G, Hamrioui B, de Lamballerie X, Charrel RN. Sandfly fever Sicilian virus, Algeria Emerg Infect Dis 2008; 14: 795-.].
Recently, the novel Massilia virus, was discovered in sandflies from southeastern France [2Charrel RN, Moureau G, Temmam S, et al. Massilia virus, a novel Phlebovirus (Bunyaviridae) isolated from sandflies in the Mediterranean Vector Borne Zoonotic Dis, Larchmont, NY 2009; 9: 519-30.].
Taken together, these recent findings led us to conduct field investigations in Algeria, a North African country where high densities of sandflies have been reported, and where they are associated with cutaneous and visceral leishmaniasis [17Dedet JP. Stages in the identification of phlebotomine sandflies as vectors of leishmaniases and other tropical diseases Parassitologia 2005; 47: 291-5., 18Harrat Z, Pratlong F, Belazzoug S, et al. Leishmania infantum and L. major in Algeria Trans R Soc Trop Med Hyg 1996; 90: 625-9.].
MATERIALS AND METHODOLOGY
Sandfly Trapping and Morphological Identification
In July 2007, sandflies were trapped in northern Algeria using previously described methodology [14Charrel RN, Izri A, Temmam S, et al. Cocirculation of 2 genotypes of Toscana virus, southeastern France Emerg Infect Dis 2007; 13: 465-8.]. Trapping was performed at two sites: (i) Larbaa Nath Iraten (previously known as Fort National) in the Kabylian region of Algeria, close to Tizi Ouzou, (ii) and Bou Ismail, in the vicinity of Alger, the capital city. CDC Miniature Light Traps were adapted to sandflies capture by using an ultra-fine mesh. Traps were placed between one and two meters above the ground. They were set during the late afternoon in the vicinity of or in animal housing facilities (sheep, goats, rabbits, chicken). Each morning, sandflies were collected, identified morphologically, and placed in 1.5mL microfuge tubes. They were organized in monospecific pools according to sex and trapping area, each pool containing up to 30 entire sandflies. Pools were individually ground in RNA Now, a chaotropic solution containing guanidinium thiocyanate (Ozyme, Montigny Le Bretonneux, France) due to a lack of dry ice or liquid nitrogen for preservation.
Virology and Phylogenetic Analysis
RNA Now-based viral RNA purification was conducted according to the manufacturer's protocol (Ozyme). A total of 10 µL of RNA was used for RT-PCR in a final volume of 50 µL as previously described [19Sanchez-Seco MP, Echevarria JM, Hernandez L, Estevez D, Navarro-Mari JM, Tenorio A. Detection and identification of Toscana and other phleboviruses by RT-nested-PCR assays with degenerated primers J Med Virol 2003; 71: 140-9.]. RT-PCR reactions were performed using the Access RT-PCR System kit (Promega France, Charbonnières). Positive PCR products of the expected size were gel-purified, cloned and sequenced in both directions. Resulting sequences were aligned together with homologous sequences of selected members of the genus Phlebovirus retrieved from Genbank. Genbank accession numbers are shown in Fig. (2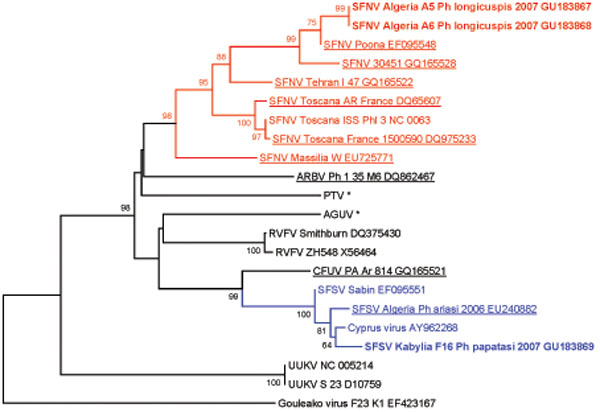 ). Sequences obtained in this study were compiled with those from Genbank to perform genetic distance comparison and phylogenetic analysis. Distance matrices were calculated for each novel sequence independently. Pairwise distance and groupings were determined via neighbor-joining method within the MEGA 4.0 software program [20Tamura K, Dudley J, Nei M, Kumar S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0 Mol Biol Evol 2007; 24: 1596-9.]. The robustness of the groupings was tested by using 1000 bootstrap pseudoreplications. Due to the preservation technique, it was impossible to attempt virus isolation on cell culture.
). Sequences obtained in this study were compiled with those from Genbank to perform genetic distance comparison and phylogenetic analysis. Distance matrices were calculated for each novel sequence independently. Pairwise distance and groupings were determined via neighbor-joining method within the MEGA 4.0 software program [20Tamura K, Dudley J, Nei M, Kumar S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0 Mol Biol Evol 2007; 24: 1596-9.]. The robustness of the groupings was tested by using 1000 bootstrap pseudoreplications. Due to the preservation technique, it was impossible to attempt virus isolation on cell culture.
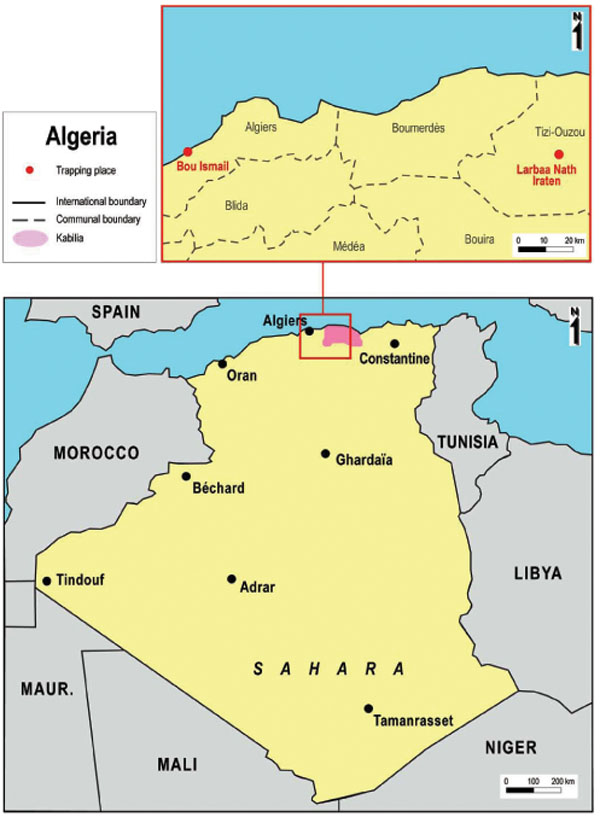 |
Fig. (1) Geographic Trapping locations in Algeria. |
 |
Fig. (2) Phylogenetic analysis of phlebovirus sequences obtained from Algerian sandflies based on 67-AA sequence representing a section of the polymerase gene. Distances and groupings were determined by the pairwise or kimura-2 algorithm and neighbor-joining method within MEGA 4.0 [20Tamura K, Dudley J, Nei M, Kumar S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0 Mol Biol Evol 2007; 24: 1596-9.]. Bootstrap values corresponding to 1000 replications are indicated. Sequences determined in this study are bolded. |
Human Sera
Two series of human sera were available for seroprevalence studies: (i) 60 samples from healthy persons living in Kabylia in the vicinity of the area where sandflies were trapped, (ii) 104 samples from patients living in the city of Alger who presented at a private biological laboratory for reasons not specifically related to febrile illness.
Serology
Immunoglobulin (Ig) G and M were detected by indirect immunofluorescence (IIF) assay as previously described [21Fulhorst CF, Monroe MC, Salas RA, et al. Isolation, characterization and geographic distribution of Cano Delgadito virus, a newly discovered South American hantavirus (family Bunyaviridae) Virus Res 1997; 51: 159-71.] with minor modifications. Briefly, equal amounts of infected and uninfected Vero cells were mixed together and spotted onto 2-well glass slides through a 3-min cytospin-based centrifugation at 900 rpm. Samples were tested at 1:20 dilution in phosphate-buffered saline. The presence of uninfected Vero cells allowed to detect non specific fluorescence (all cells were fluorescent). Sera were tested by IgG and IgM enzyme-linked immunosorbent assay (ELISA) with the Toscana virus - Enzywell Toscana virus (recombinant nucleoprotein) IgG and IgM - DIESSE Diagnostica Senese S.p.A, (Siena, Italy) using the protocol recommended by the manufacturer. A serum was considered positive in either IgG or IgM if both ELISA and IIF were positive.
RESULTS
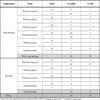
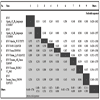
The two trapping sites are shown in Fig. (1 ). A total of 785 sandflies were trapped, processed individually for morphological identification, organized into 40 pools, and used for viral RNA detection (Table 1). A total of 3 pools tested positive with nested primers NPhlebo2+ / NPhlebo2-. A 201-nt sequence (excluding primers) was obtained. These 3 sequences were deposited in Genbank with accession numbers GU183867-183869. Nucleotide and amino acid distances are presented in Table 2. The two sequences obtained from Phlebotomus longicuspis were most closely related to the Poona strain of SFNV, and clearly belonged to the species SFNV, with an amino acid and nucleotide distance comprised between 9 to 37.3% and 1 to 34.8% respectively. The sequence determined from Phlebotomus papatasi was most closely related to Cyprus virus and not to the SFSV-like phlebovirus previously discovered in Algeria in 2006. However, these 4 sequences formed a cluster together with the sequence of the prototype Sabin strain of SFSV, with pairwise amino acid and nucleotide distances lower than 6% and 20%, respectively.
). A total of 785 sandflies were trapped, processed individually for morphological identification, organized into 40 pools, and used for viral RNA detection (Table 1). A total of 3 pools tested positive with nested primers NPhlebo2+ / NPhlebo2-. A 201-nt sequence (excluding primers) was obtained. These 3 sequences were deposited in Genbank with accession numbers GU183867-183869. Nucleotide and amino acid distances are presented in Table 2. The two sequences obtained from Phlebotomus longicuspis were most closely related to the Poona strain of SFNV, and clearly belonged to the species SFNV, with an amino acid and nucleotide distance comprised between 9 to 37.3% and 1 to 34.8% respectively. The sequence determined from Phlebotomus papatasi was most closely related to Cyprus virus and not to the SFSV-like phlebovirus previously discovered in Algeria in 2006. However, these 4 sequences formed a cluster together with the sequence of the prototype Sabin strain of SFSV, with pairwise amino acid and nucleotide distances lower than 6% and 20%, respectively.

The two seroprevalence studies indicated that the presence of IgG specific of SFNV or SFN-like virus was observed at rates of 10.6% (11/104 sera) and 21.6% (13/60 sera) originating from Bou Ismail and Larbaa Nath Iraten areas, respectively. In the Bou Ismail subset, the mean age of seropositives was 49 while the mean age of the total population was 37.
DISCUSSION
Recent investigations of sandfly-transmitted phleboviruses have indicated that virus diversity within the Mediterranean is higher than initially suspected, and that local populations have a high risk of infection during their lifetime [2Charrel RN, Moureau G, Temmam S, et al. Massilia virus, a novel Phlebovirus (Bunyaviridae) isolated from sandflies in the Mediterranean Vector Borne Zoonotic Dis, Larchmont, NY 2009; 9: 519-30., 3Izri A, Temmam S, Moureau G, Hamrioui B, de Lamballerie X, Charrel RN. Sandfly fever Sicilian virus, Algeria Emerg Infect Dis 2008; 14: 795-., 14Charrel RN, Izri A, Temmam S, et al. Cocirculation of 2 genotypes of Toscana virus, southeastern France Emerg Infect Dis 2007; 13: 465-8., 22Charrel RN, Gallian P, Navarro-Mari JM, et al. Emergence of Toscana virus in Europe Emerg Infect Dis 2005; 11: 1657-63.-24Sanbonmatsu-Gamez S, Perez-Ruiz M, Collao X, et al. Toscana virus in Spain Emerg Infect Dis 2005; 11: 1701-7.]. These recent data suggest that sandfly associated viral infections may also be a concern for public health in regions located south of the Mediterranean, where high densities of sandflies are reported. For these reasons, we decided to conduct investigations based on a transversal approach involving at first a combination of entomological and virological studies, and seroprevalence studies on populations living in the area where sandflies were trapped. A total of one and two pools were found to contain SFS- and SFN-like virus RNA, respectively. Human infection due to these viruses was supported by seroprevalence studies indicating rates varying from 5% for SFS-like virus [3Izri A, Temmam S, Moureau G, Hamrioui B, de Lamballerie X, Charrel RN. Sandfly fever Sicilian virus, Algeria Emerg Infect Dis 2008; 14: 795-.] and 10.6-21.6% for SFN-like virus (this study). The new SFS-like virus was detected in Phlebotomus papatasi and showed to be closely related to, but distinct from, SFSV previously detected in Phlebotomus ariasi from Algeria [3Izri A, Temmam S, Moureau G, Hamrioui B, de Lamballerie X, Charrel RN. Sandfly fever Sicilian virus, Algeria Emerg Infect Dis 2008; 14: 795-.], and from Cyprus virus isolated from humans [23Papa A, Konstantinou G, Pavlidou V, Antoniadis A. Sandfly fever virus outbreak in Cyprus Clin Microbiol Infect 2006; 12: 192-4.-25Konstantinou GN, Papa A, Antoniadis A. Sandfly-fever outbreak in Cyprus: are phleboviruses still a health problem? Travel Med Infect Dis 2007; 5: 239-42.].
Since detection of viral sequences could not be confirmed through virus isolation, it is necessary to address the question of potential contamination. Concerning the two SFN-like phleboviruses, the sequences are genetically sufficiently distinct from other sequences determined in our unit during the last 3 years (underlined in the Fig. 2) to exclude laboratory contamination. Moreover, the genetic variation inside the Sandfly fever Naples group is much larger than determined in our previous study [2Charrel RN, Moureau G, Temmam S, et al. Massilia virus, a novel Phlebovirus (Bunyaviridae) isolated from sandflies in the Mediterranean Vector Borne Zoonotic Dis, Larchmont, NY 2009; 9: 519-30.]. Based on the polymerase gene, the genetic distance between this newly discovered strain of Sandfly fever Naples in Algeria and Massilia virus is 37.3% and 34.3% for amino acids and nucleotide data respectively. The novel SFSV-like sequence obtained in this study from the Phlebotomus papatasi pool is genetically divergent at 4 nt sites and 4 AA sites from sequences previously determined from a pool of Phlebotomuspatatasi in our laboratory in 2007 [3Izri A, Temmam S, Moureau G, Hamrioui B, de Lamballerie X, Charrel RN. Sandfly fever Sicilian virus, Algeria Emerg Infect Dis 2008; 14: 795-.] and at 2 nt sites and 2 AA sites from Sandfly fever Sicilian strain Sabin. Furthermore, detailed analysis of the relevant chromatograms supports this clear divergence.
SFSV and SFS-Like Viruses
Human cases of SFSV infection have been reported from Italy, Egypt, Pakistan, Iran and Cyprus [16Karabatsos N. International Catalogue of Arthropod-borne viruses In: American Society for Tropical Medicine and Hygiene. 3rd. San Antonio, Texas 1985; (Suppl 1): pp. 137-52., 23Papa A, Konstantinou G, Pavlidou V, Antoniadis A. Sandfly fever virus outbreak in Cyprus Clin Microbiol Infect 2006; 12: 192-4.]. Seroprevalence studies performed with human or vertebrate sera indicates that SFSV, or a closely related one, is circulating in Jordan [26Batieha A, Saliba EK, Graham R, Mohareb E, Hijazi Y, Wijeyaratne P. Seroprevalence of West Nile, Rift Valley, and sandfly arboviruses in Hashimiah, Jordan Emerg Infect Dis 2000; 6: 358-62.], Israel [27Cohen D, Zaide Y, Karasenty E, et al. Prevalence of antibodies to West Nile fever, sandfly fever Sicilian, and sandfly fever Naples viruses in healthy adults in Israel Public Health Rev 1999; 27: 217-30.], Sudan [28McCarthy MC, Haberberger RL, Salib AW, et al. Evaluation of arthropod-borne viruses and other infectious disease pathogens as the causes of febrile illnesses in the Khartoum Province of Sudan J Med Virol 1996; 48: 141-6.], Tunisia [29Chastel C, Bach-Hamba D, Launay H, Le Lay G, Hellal H, Beaucournu JC. [Arbovirus infections in Tunisia: new serological survey of small wild mammals] Bull Soc Pathol Exot Filiales 1983; 76: 21-33.], Pakistan [30Darwish MA, Hoogstraal H, Roberts TJ, Ghazi R, Amer T. A sero-epidemiological survey for Bunyaviridae and certain other arboviruses in Pakistan Trans R Soc Trop Med Hyg 1983; 77: 446-50.], Egypt [31Darwish MA, Feinsod FM, Scott RM, et al. Arboviral causes of non-specific fever and myalgia in a fever hospital patient population in Cairo, Egypt Trans R Soc Trop Med Hyg 1987; 81: 1001-3.], Bangladesh [32Gaidamovich SY, Baten MA, Klisenko GA, Melnikova YE. Serological studies on sandfly fevers in the Republic of Bangladesh Acta Virol 1984; 28: 325-8.], Iran [33Saidi S, Tesh R, Javadian E, Sahabi Z, Nadim A. Studies on the epidemiology of sandfly fever in Iran. II. The prevalence of human and animal infection with five phlebotomus fever virus serotypes in Isfahan province Am J Trop Med Hyg 1977; 26: 288-93.]. At the outset of this study, the most comprehensive serological study did not detect the presence of neutralizing antibodies reactive to SFSV in human sera of Algerian populations of Tamanrasset and Djanet [34Tesh RB, Saidi S, Gajdamovic SJ, Rodhain F, Vesenjak-Hirjan J. Serological studies on the epidemiology of sandfly fever in the Old World Bull World Health Organ 1976; 54: 663-74.]. However, these two areas are located at the southern border of Algeria in arid environment. Detection of SFS-like RNA in this study corroborates previous findings concerning the presence of a new phlebovirus closely related to but distinct from SFSV and Cyprus virus. It is likely that the Algerian SFS-like virus infects humans according to serology results previously reported [3Izri A, Temmam S, Moureau G, Hamrioui B, de Lamballerie X, Charrel RN. Sandfly fever Sicilian virus, Algeria Emerg Infect Dis 2008; 14: 795-.].
SFNV and SFN-Like Viruses
Human cases of SFNV, mostly caused by Toscana virus, based on virus isolation and/or molecular evidence have been reported from North Mediterranean counties [6Valassina M, Meacci F, Valensin PE, Cusi MG. Detection of neurotropic viruses circulating in Tuscany: the incisive role of Toscana virus J Med Virol 2000; 60: 86-90., 13Peyrefitte CN, Devetakov I, Pastorino B, et al. Toscana virus and acute meningitis, France Emerg Infect Dis 2005; 11: 778-80.-15Santos L, Simoes J, Costa R, Martins S, Lecour H. Toscana virus meningitis in Portugal, 2002-2005 Euro Surveill 2007; 12: E3-4., 24Sanbonmatsu-Gamez S, Perez-Ruiz M, Collao X, et al. Toscana virus in Spain Emerg Infect Dis 2005; 11: 1701-7., 35Baldelli F, Ciufolini MG, Francisci D, et al. Unusual presentation of life-threatening Toscana virus meningoencephalitis Clin Infect Dis 2004; 38: 515-20.-38Valassina M, Valentini M, Valensin PE, Cusi MG. Fast duplex one-step RT-PCR for rapid differential diagnosis of entero- or toscana virus meningitis Diagn Microbiol Infect Dis 2002; 43: 201-5.]. The original Naples virus was identified for the last time in 1985 in Egypt from a human specimen [39Liu DY, Tesh RB, Travassos Da Rosa AP, et al. Phylogenetic relationships among members of the genus Phlebovirus (Bunyaviridae) based on partial M segment sequence analyses J Gen Virol 2003; 84: 465-73.]. For the last two decades, Toscana virus was the only genotype of the SFNV species to have been detected and/or isolated in relation to human cases [22Charrel RN, Gallian P, Navarro-Mari JM, et al. Emergence of Toscana virus in Europe Emerg Infect Dis 2005; 11: 1657-63.]. Seroprevalence studies performed with human sera indicate that SFNV, or a closely related virus, is circulating in Ethiopia, Sudan, Egypt, Morocco, Greece, former Yugoslavia, Turkey, Iraq, Saudi Arabia, Iran, former USSR [Moldavia, Azerbaijan, Uzbekistan, Tajikistan, Turkmenistan], Pakistan and Bangladesh [34Tesh RB, Saidi S, Gajdamovic SJ, Rodhain F, Vesenjak-Hirjan J. Serological studies on the epidemiology of sandfly fever in the Old World Bull World Health Organ 1976; 54: 663-74.]. More recently, seroprevalence studies indicated that Toscana virus, or an antigenically closely-related virus, circulates in Spain [9Mendoza-Montero J, Gamez-Rueda MI, Navarro-Mari JM, de la Rosa-Fraile M, Oyonarte-Gomez S. Infections due to sandfly fever virus serotype Toscana in Spain Clin Infect Dis 1998; 27: 434-6.], France [40De Lamballerie X, Tolou H, Durand JP, Charrel RN. Prevalence of Toscana virus antibodies in volunteer blood donors and patients with central nervous system infections in southeastern France Vector borne Zoonotic Dis Larchmont NY 2007; 7: 275-7.], Italy [38Valassina M, Valentini M, Valensin PE, Cusi MG. Fast duplex one-step RT-PCR for rapid differential diagnosis of entero- or toscana virus meningitis Diagn Microbiol Infect Dis 2002; 43: 201-5., 41Francisci D, Papili R, Camanni G, et al. Evidence of Toscana virus circulation in Umbria: first report Eur J Epidemiol 2003; 18: 457-9.], and Cyprus [42Eitrem R, Stylianou M, Niklasson B. High prevalence rates of antibody to three sandfly fever viruses (Sicilian, Naples, and Toscana) among Cypriots Epidemiol Infect 1991; 107: 685-91.]. In contrast, low prevalence of antibody was observed in Germany [43Schwarz TF, Jager G, Gilch S, Pauli C. Serosurvey and laboratory diagnosis of imported sandfly fever virus, serotype Toscana, infection in Germany Epidemiol Infect 1995; 114: 501-10.]. The most comprehensive serological study did not detect the presence of neutralizing antibodies reactive to SFNV or SFN-like virus in human sera of Algerian populations of Tamanrasset and Djanet [34Tesh RB, Saidi S, Gajdamovic SJ, Rodhain F, Vesenjak-Hirjan J. Serological studies on the epidemiology of sandfly fever in the Old World Bull World Health Organ 1976; 54: 663-74.]. Therefore, at the outset of this study, there was neither molecular evidence nor serology-based data suggesting the presence of SFNV in Algeria. To our knowledge, this is the first time (i) that SFN-like virus has been detected in Algeria, (ii) and that indirect evidence via seroprevalence studies, is reported. Discrepancy between the mean age of the tested population and that of seropositive individuals is indicative that the longer the exposure, the higher the risk of getting infected by SFNV or SFN-like virus.
Together, molecular and serological data provided here constitute evidence that novel phleboviruses distinct from but related to SFSV and SFNV are present in Algeria, and can infect humans.
ACKNOWLEDGEMENTS
This work was partially supported by the French Research Agency (ANR) through the PhleboMED project, the FP7 European project n°228292 EVA (European Virus Archive). The authors thank Fabienne Pinna for preparing the immunofluorescence slides.