- Home
- About Journals
-
Information for Authors/ReviewersEditorial Policies
Publication Fee
Publication Cycle - Process Flowchart
Online Manuscript Submission and Tracking System
Publishing Ethics and Rectitude
Authorship
Author Benefits
Reviewer Guidelines
Guest Editor Guidelines
Peer Review Workflow
Quick Track Option
Copyediting Services
Bentham Open Membership
Bentham Open Advisory Board
Archiving Policies
Fabricating and Stating False Information
Post Publication Discussions and Corrections
Editorial Management
Advertise With Us
Funding Agencies
Rate List
Kudos
General FAQs
Special Fee Waivers and Discounts
- Contact
- Help
- About Us
- Search


The Open Paleontology Journal
(Discontinued)
ISSN: 1874-4257 ― Volume 5, 2014
Timing of Shell Size Increase and Decrease of the Planktic Foraminifer Neogloboquadrina pachyderma (Sinistral) During the Pleistocene, IODP Exp. 303 Site U1304, the North Atlantic Ocean
M Yamasaki*, 1, M Matsui1, C. Shimada2, S. Chiyonobu3, T. Sato1
Abstract
We made shell size measurements of the planktic foraminifer Neogloboquadrina pachyderma (sinistral) using samples from IODP Site U1304, the North Atlantic. We found that mean and maximum shell sizes began to increase around 1.1 Ma, and that several episodic changes in test size occurred during 0.6 – 0.35 Ma. Test size reached a maximum during the late Quaternary. Based on correlation with previous investigations, we have confirmed that these changes in foraminiferal shell size occurred on an inter-ocean scale
Article Information
Identifiers and Pagination:
Year: 2008Volume: 1
First Page: 18
Last Page: 23
Publisher Id: TOPALOJ-1-18
DOI: 10.2174/1874425700801010018
Article History:
Received Date: 11/5/2008Revision Received Date: 1/8/2008
Acceptance Date: 1/8/2008
Electronic publication date: 28/08/2008
Collection year: 2008
open-access license: This is an open access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.5/), which permits unrestrictive use, distribution, and reproduction in any medium, provided the original work is properly cited.
* Address correspondence to this author at the Institute of Applied Earth Sciences, Faculty of Engineering and Resource Science, Akita University, Akita 010-8502, Japan; Fax: +81-18-837-0401; E-mail: yamasaki@ipc.akita-u.ac.jp
| Open Peer Review Details | |||
|---|---|---|---|
| Manuscript submitted on 11-5-2008 |
Original Manuscript | Timing of Shell Size Increase and Decrease of the Planktic Foraminifer Neogloboquadrina pachyderma (Sinistral) During the Pleistocene, IODP Exp. 303 Site U1304, the North Atlantic Ocean | |
INTRODUCTION
High latitude boreal seas are characterized by the extraordinary abundance of the planktic foraminifer Neogloboquadrina pachyderma (sinistral) (e.g. [1, 2]). Shell size of this species has recently been reported to have increased through the Quaternary in northern hemisphere oceans [3, 4]. Kucera and Kennett [4] found such trend in the North Pacific and claimed that “the modern type” of the species had appeared about 1 Ma. Generally, the shell size of foraminifera is believed to have increased during the Cenozoic, and maximized in the most favorable conditions [5]. In the case of N. pachyderma (sin), Huber et al. [3] suggested that such an increase in shell size would reflect their evolutional adaptation when cold waters progressively appeared in the high latitudinal seas together with increasing glaciation since the mid-Quaternary. However, most of these previous works have targeted upper Pleistocene and Holocene (1.3 Ma to the Recent) deposits off California and the Norwegian - Greenland Sea. In both regions N. pachyderma (sin) is not frequent but rather of sporadic occurrence during the early Pleistocene. In this paper, we describe the long-term trend in shell size change of N. pachyderma (sin) from its main habitat, and to investigate the detailed timing of size increase using good quality core materials covering of the whole Quaternary period in the subarctic North Atlantic.
MATERIALS AND METHODOLOGY
Materials were obtained from IODP Site U1304 (53˚03’N, 33˚31’W; water depth, 3080 m), located within the North Atlantic subarctic convergence zone (Fig. 1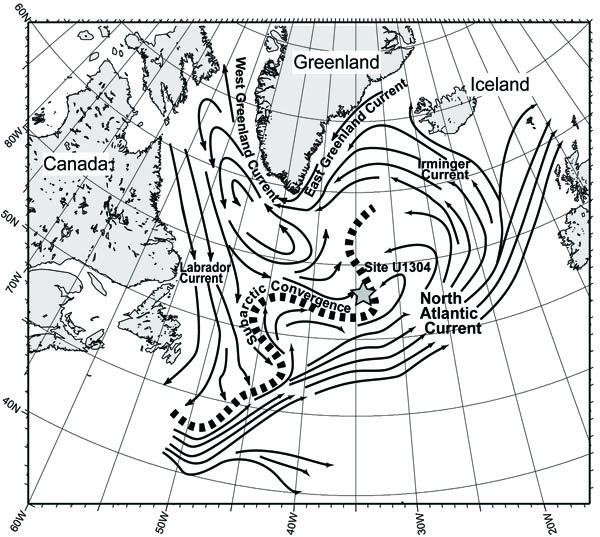 ). The sediments at the site consist of biogenic components, alternation calcareous nannofossil ooze and laminated diatomaceous ooze, with less common intervals of clay and silty clay which is rich in microfossils [6].
). The sediments at the site consist of biogenic components, alternation calcareous nannofossil ooze and laminated diatomaceous ooze, with less common intervals of clay and silty clay which is rich in microfossils [6].
 |
Fig. (1) Map showing the surface ocean currents, the North Atlantic subarctic convergence zone [21], and the study location. |
An age model for Site U1304 was established using paleomagnetostratigraphy and biostratigraphy of both calcareous nannofossil and diatom [6]. In total, 16 biostratigraphic datum levels spanning the last 1.8 million years were employed in this study.
For foraminiferal analyses including size measurements, 89 horizons were chosen down to 273.09 meters composite depth (mcd) from Holes U1304A and U1304B. Each 10 cm3 sample was first soaked in a 3 % hydrogen peroxide solution for a few hours. Samples were then washed through a 63-µm screen, and the retained particles were dried at 60˚C. The size fraction larger than 63 µm was split into smaller aliquots with a microsplitter. Each sample was then dry sieved through a 125-µm screen. Planktic foraminiferal shells larger than 125 µm were picked from the aliquots until ideally 200 specimens were collected. All specimens were identified and counted. All specimens of N. pachyderma (sin) were oriented in the standard taxonomic umbilical view, and the maximum diameter of each specimen was measured and given as “shell size”, using the public domain image analyzing software ImageJ (http://rsb.info.nih.gov/ij/index.html). In the case of samples with very low N. pachyderma (sin) abundances, another aliquot was used for obtaining additional specimens of N. pachyderma (sin) for the size measurement. In our analysis, both of the two distinct ultrastructural morphotypes of N. pachyderma (sin) were measured, (1) the crystalline and (2) the reticulate morphotypes [7]. In total, 8849 specimens were measured. The mean, the 25 and 75 percentiles, outliers, and the range, which shows an interval between the 25 and 75 percentile, are given in Fig. (2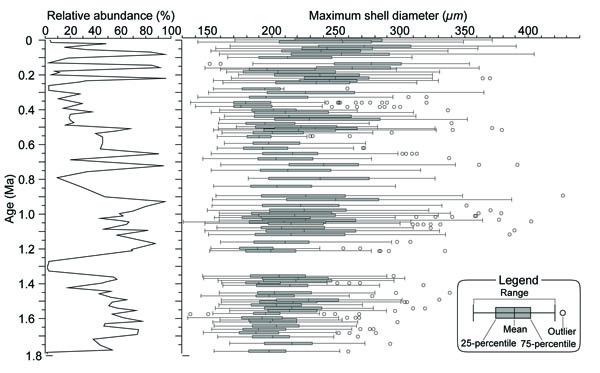 ).
).

RESULTS
Neogloboquadrina pachyderma (sin) was usually the most dominant species in the samples analysed (Table 1), accounting for up to 96.2% in relative abundance and 42.5% on average of the total planktic foraminiferal assemblages (Fig. 2). N. pachyderma (sin) increased in particular from the lowermost Pleistocene, confirming the work of Weaver and Clement [8] who presented a framework of the planktic foraminiferal biostratigraphy in the North Atlantic, and is also consistent with the report of Meggers and Baumann [9], who showed that N. pachyderma (sin) abundantly occurred in the Labrador Sea since 1.1 Ma. The abundance of N. pachyderma (sin) dropped to <50% around 1.3 and 0.8 Ma, and from ca. 0.6 to 0.25 Ma.

The mean shell size (MSS) from a total of 8849 specimens was 220.6 µm. In addition, the MSS ranged from 187.0 to 237.7 µm while sizes as large as 400 µm were recorded in some horizons (Fig. 2 and Table 1). Both MSS and range in shell size were generally low (190.9 – 227.5 µm) between 1.8 and 1.2 Ma. On the other hand, larger shells were often observed between 1.1 – 0.9 Ma, with a 75 percentile larger than 250 µm. At the same time, ranges became wider than that of the lower. Smaller values of MSS were recognizable again between 0.6 – 0.35 Ma, and the minimum MSS was 187.0 µm at 0.36 Ma. In the following, there was a significant increase of MSS, and MSS eventually became larger than 250 µm from 0.35 Ma to the Recent. Though the correlation between MSS and percent abundance of N. pachyderma (sin) is low (r=0.19), smaller MSS were observed at low abundances of N. pachyderma (sin) within the total planktic foraminiferal assemblage during some periods (e.g., around 0.4 Ma).
DISCUSSION
Shell size changes of N. pachyderma (sin) during the late Quaternary were previously reported from the Norwegian – Greenland Sea [3] and the North Pacific [4]. We can directly correlate our dataset with these studies, comparing the maximum diameter of N. pachyderma (sin). The study area is very appropriate for obtaining a long-term record of morphological change in N. pachyderma (sin), because environmental conditions at the southernmost part of the subarctic water mass potentially provide robust occurrences of the species during the Quaternary. In fact, N. pachyderma (sin) has continuously occurred in moderate to high abundance throughout the Quaternary section at Site U1304 (Table 1).
Based on the temporal changes of MSS and occurrences of large sized specimens in our study, N. pachyderma (sin) clearly shows a remarkable variation in shell size after 1.1 Ma (Fig. 2). Especially, larger specimens (>350 µm) occur in the interval following 1.1 Ma. The temporal size changes are consistent with the previously obtained data from the Norwegian – Greenland Sea at 1.1 – 1.0 Ma [3] and the northeastern Pacific around 1 Ma [4]. Thus, the shell size change around 1.1 Ma is a bio-event on an inter-ocean basin scale. The correlation between Site U1304 and the other sites [3, 4] previously studied in terms of MSS changes clearly shows that MSS at all sites increased throughout the late Quaternary (Fig. 3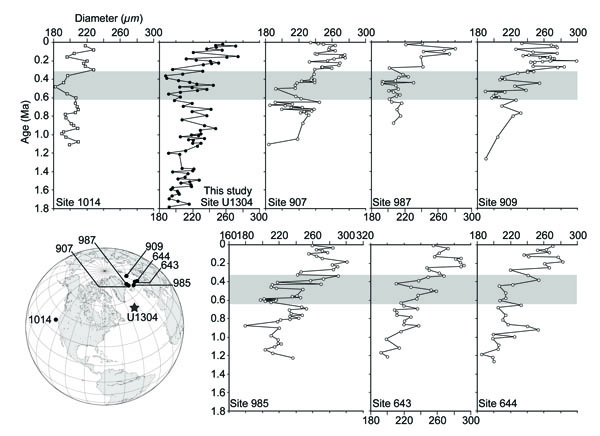 ). Evidence presented by Huber et al. [3] indicates that the largest shell size occurred after 0.4 Ma in the Norwegian – Greenland Sea, confirmed by our data (Fig. 2). The MSS change at Site U1304 is further characterized by a few episodic decreases and increases between 0.6 – 0.35 Ma, a similar trend to that observed from Site 1014 in the North Pacific [4]. The minimum value of MSS was found in this interval at Sites U1304 and ODP Site 1014. In this interval, the shell size recessed its trend of increasing. These falloff events also occurred at Sites 907 and 987, in the western Norwegian – Greenland Sea, and Site 909, in the Fram Strait, especially onset of that event is ca. 0.6 Ma. On the other hand, these events were not clear in the eastern Norwegian – Greenland Sea (Sites 985, 643 and 644). In those three sites, the shell size seems to increase rather than decrease between 0.6 – 0.35 Ma although there are some episodic decrease.
). Evidence presented by Huber et al. [3] indicates that the largest shell size occurred after 0.4 Ma in the Norwegian – Greenland Sea, confirmed by our data (Fig. 2). The MSS change at Site U1304 is further characterized by a few episodic decreases and increases between 0.6 – 0.35 Ma, a similar trend to that observed from Site 1014 in the North Pacific [4]. The minimum value of MSS was found in this interval at Sites U1304 and ODP Site 1014. In this interval, the shell size recessed its trend of increasing. These falloff events also occurred at Sites 907 and 987, in the western Norwegian – Greenland Sea, and Site 909, in the Fram Strait, especially onset of that event is ca. 0.6 Ma. On the other hand, these events were not clear in the eastern Norwegian – Greenland Sea (Sites 985, 643 and 644). In those three sites, the shell size seems to increase rather than decrease between 0.6 – 0.35 Ma although there are some episodic decrease.

Size changes of planktic foraminifers are believed to be caused by environmental change. Planktic foraminifers typically reach their maximum size within the most favorable environmental conditions, and decrease in size away from such areas [10, 11]. The period around ca. 0.4 Ma is recognized as “the mid-Brunhes event (MBE)”, characterized by more humid interglacial climates with long-lasting warmth [12, 13] and also by an unusual global increase in pelagic carbonate production and dissolution [14]. Some unusual sedimentary facies such as laminated diatomaceous oozes as well as hiatuses, were found at around 0.54 -0.53 Ma in southern Atlantic pelagic sediments, and these events were considered to be one of the terminal events of the Mid-Pleistocene climatic Transition (MPT; [15]). Furthermore, Wright and Flower [16] suggested that surface water conditions had changed in the subarctic North Atlantic from about 0.66 – 0.61 Ma, coinciding with the establishment of “the 100-kyr world”. The intervals of N. pachyderma (sin) shell size change correspond to the timing between the end of the MPT and the MBE. We thus suggest that the predominance of humid and long-lasting warmer conditions and/or surface water instability at about 0.6 – 0.35 Ma was likely to prevent faunal dominance of N. pachyderma (sin), and resulting in the decrease of the shell size. These morphological findings confirm the adaptation of this species to cold water habitats.
Recently, molecular biological evidence has revealed the phylogenetic origin of this species [17, 18, 19]. Darling et al. [20] suggested that populations of N. pachyderma (sin) between the North Atlantic and North Pacific oceans have been genetically isolated from each other with no gene flow since the early Pleistocene. If this is true, the shell size change of N. pachyderma (sin) during ca. 0.6 - 0.35 Ma can be interpreted as a sensitive reaction of the species to environmental and climate change common at both oceans the North Atlantic and eastern North Pacific.
CONCLUSIONS
Neogloboquadrina pachyderma (sin) has increased in mean test size since the mid-Quaternary (1.1 Ma). This trend in test size at the North Atlantic subarctic convergence zone is consistent with test size changes reported from other regions of the northern Atlantic [3] and the eastern Pacific [4]. Based on a correlation of shell size change of this species in the North Atlantic and North Pacific, shell size decrease event around ca. 0.6 Ma is supposed to be an inter-ocean scaled bioevent, caused by changes of the global climate.
ACKNOWLEDGEMENTS
We sincerely appreciate the support of captain, crews and technical staffs of IODP Expedition 303. Thanks also to Saiko Toyoshima for micropaleontological sample preparations. R. Schiebel at University of Southamptonand two anonymous reviewers helped to improve the manuscript with constructive comments. Financial support was given to TS (Grand-In –Aid for Scientific Research; No. 19340155).