- Home
- About Journals
-
Information for Authors/ReviewersEditorial Policies
Publication Fee
Publication Cycle - Process Flowchart
Online Manuscript Submission and Tracking System
Publishing Ethics and Rectitude
Authorship
Author Benefits
Reviewer Guidelines
Guest Editor Guidelines
Peer Review Workflow
Quick Track Option
Copyediting Services
Bentham Open Membership
Bentham Open Advisory Board
Archiving Policies
Fabricating and Stating False Information
Post Publication Discussions and Corrections
Editorial Management
Advertise With Us
Funding Agencies
Rate List
Kudos
General FAQs
Special Fee Waivers and Discounts
- Contact
- Help
- About Us
- Search




The Open Cancer Immunology Journal
(Discontinued)
ISSN: 1876-4010 ― Volume 8, 2020
The Antitumor Immunity Mediated by NK Cells: The Role of The NCRs
Mona Rady*, Khaled Abou-Aisha
Abstract
Natural Killer (NK) cells are innate immune lymphocytes that are important for early and effective immune responses against infections and cancer. The antitumor immunity mediated by NK cells can be exerted through several direct or indirect “immunosurveillance” mechanisms that control tumor growth and prevent the rapid dissemination of metastatic tumors. NK cells express an array of activating and inhibitory receptors that enable them to recognize and bind non-self as well as self-ligands expressed on the surface of malignant or virally infected cells. The family of Natural Cytotoxicity Receptors (NCRs) comprises three activating receptors; NKp30, NKp44, and NKp46 that are important for the stimulation of NK cell effector functions. This review summarizes the mechanisms of antitumor immunity mediated by natural killer cells with focus on the role of the family of the NCRs and their tumor associated ligands.
Article Information

Identifiers and Pagination:
Year: 2018Volume: 07
First Page: 7
Last Page: 15
Publisher Id: TOCIJ-7-7
DOI: 10.2174/1876401001807010007
Article History:
Received Date: 31/12/2017Revision Received Date: 10/05/2018
Acceptance Date: 29/05/2018
Electronic publication date: 20/06/2018
Collection year: 2018
open-access license: This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International Public License (CC-BY 4.0), a copy of which is available at: (https://creativecommons.org/licenses/by/4.0/legalcode). This license permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
* Address correspondence to this author at the Microbiology and Immunology Department, German University in Cairo (GUC), New Cairo, Egypt, Tel: +201005633776; E-mails: mona.rady@gmail.com; mona.rady@guc.edu.eg
| Open Peer Review Details | |||
|---|---|---|---|
| Manuscript submitted on 31-12-2017 |
Original Manuscript | The Antitumor Immunity Mediated by NK Cells: The Role of The NCRs | |
1. INTRODUCTION
Historically, the histologic and functional definition of a natural killer cell (NK cell) was that of a Large Granular Lymphocyte (LGL) that could kill a target cell “naturally”, that is, spontaneously without any prior sensitization and without restriction by the Major Histocompatibility Complex (MHC) [1Herberman RB, Nunn ME, Lavrin DH. Natural cytotoxic reactivity of mouse lymphoid cells against syngeneic acid allogeneic tumors. I. Distribution of reactivity and specificity. Int J Cancer 1975; 16(2): 216-29.
[http://dx.doi.org/10.1002/ijc.2910160204] [PMID: 50294] , 2Kiessling R, Klein E, Pross H, Wigzell H. Natural killer cells in the mouse. II. Cytotoxic cells with specificity for mouse Moloney leukemia cells. Characteristics of the killer cell. Eur J Immunol 1975; 5(2): 117-21.
[http://dx.doi.org/10.1002/eji.1830050209] [PMID: 1086218] ]. NK cells are effector lymphocytes that belong to the innate immune system and exert multiple functions among which the rapid elimination of virus-infected or transformed cells is the most important. NK cells are regarded as innate immune lymphocytes because unlike T and B lymphocytes, NK cells’ effector functions are controlled by a repertoire of germline-encoded receptors that do not undergo somatic recombination [3Cheng M, Chen Y, Xiao W, Sun R, Tian Z. NK cell-based immunotherapy for malignant diseases. Cell Mol Immunol 2013; 10(3): 230-52.
[http://dx.doi.org/10.1038/cmi.2013.10] [PMID: 23604045] ]. Despite these known innate immune cell functions, there is some evidence that NK cells possess some features as an “adaptive” immune component as well. Accumulating evidences suggest that NK possess receptors with some antigen specificity and can in fact undergo clonal expansion following antigen exposure. Moreover, NK cells can “remember” the antigen; a phenomenon only characteristic of adaptive immune components, generating long-lived memory cells [4Paust S, von Andrian UH. Natural killer cell memory. Nat Immunol 2011; 12(6): 500-8.
[http://dx.doi.org/10.1038/ni.2032] [PMID: 21739673] -6Vivier E, Raulet DH, Moretta A, et al. Innate or adaptive immunity? The example of natural killer cells. Science 2011; 331(6013): 44-9.
[http://dx.doi.org/10.1126/science.1198687] [PMID: 21212348] ]. NK cells develop in Bone Marrow (BM) from common lymphoid progenitor cells. After development, NK cells distribute widely throughout various lymphoid tissues other than BM, such Lymph Nodes (LN) and spleen as well as non-lymphoid tissues such as peripheral blood, lung and liver [5Sun JC, Lanier LL. NK cell development, homeostasis and function: Parallels with CD8+ T cells. Nat Rev Immunol 2011; 11(10): 645-57.
[http://dx.doi.org/10.1038/nri3044] [PMID: 21869816] ]. The traditional cell surface phenotype of human NK cells is defined by absence of CD3 (thereby excluding T cells) and expression of CD56, the 140-kDa isoform of Neural Cell Adhesion Molecule (NCAM) [7Lanier LL, Testi R, Bindl J, Phillips JH. Identity of Leu-19 (CD56) leukocyte differentiation antigen and neural cell adhesion molecule. J Exp Med 1989; 169(6): 2233-8.
[http://dx.doi.org/10.1084/jem.169.6.2233] [PMID: 2471777] , 8Ritz J, Schmidt RE, Michon J, Hercend T, Schlossman SF. Characterization of functional surface structures on human natural killer cells. Adv Immunol 1988; 42: 181-211.
[http://dx.doi.org/10.1016/S0065-2776(08)60845-7] [PMID: 3284289] ]. NK cells represent 2% to18% of total lymphocytes in human peripheral blood. In humans, NK cells can be divided into CD56dim and CD56bright NK cell subsets based on levels of cell surface expression of CD56. CD56dim NK cells are the predominant NK subset in peripheral blood, while CD56bright NK cells which are more abundant in lymphoid organs [9Vivier E, Tomasello E, Baratin M, Walzer T, Ugolini S. Functions of natural killer cells. Nat Immunol 2008; 9(5): 503-10.
[http://dx.doi.org/10.1038/ni1582] [PMID: 18425107] ].
2. ANTITUMOR IMMUNITY MEDIATED BY NK CELLS
The antitumor immunity mediated by NK cells can be exerted through several direct or indirect “immunosurveillance” mechanisms. By infiltrating through the tumor microenvironment, NK cells recognize, attack and kill tumor cells, there by controlling the growth of primary tumors. NK cells also prevent the rapid dissemination of metastatic tumors by killing circulating cancer cells that metastasize from the tumour microenvironment to other organs. NK cells can directly kill target tumor cells by releasing cytoplasmic granules containing perforin (membrane pore-forming proteins) and granzymes (serine proteases) that leads to tumor-cell apoptosis (programmed cell death) by caspase-dependent and -independent pathways [10Kägi D, Ledermann B, Bürki K, et al. Cytotoxicity mediated by T cells and natural killer cells is greatly impaired in perforin-deficient mice. Nature 1994; 369(6475): 31-7.
[http://dx.doi.org/10.1038/369031a0] [PMID: 8164737] , 11Trapani JA, Davis J, Sutton VR, Smyth MJ. Proapoptotic functions of cytotoxic lymphocyte granule constituents in vitro and in vivo. Curr Opin Immunol 2000; 12(3): 323-9.
[http://dx.doi.org/10.1016/S0952-7915(00)00094-7] [PMID: 10781403] ]. Cytotoxic granules reorient towards the tumor cell soon after NK–tumor cell interaction and are released into the immunological synapse in a calcium-dependent manner. Perforin-induced membrane perforations allow the delivery of granzymes into tumor cells, leading to induction of apoptosis. Another mechanism by which NK cells directly kill tumor cells is death receptor-mediated apoptosis when FasL or TNF-Related Apoptosis-Inducing Ligand (TRAIL) expressed on NK cells binds to their respective target cell surface death receptors; Fas and TRAIL receptor (TRAILR) [12Zamai L, Ahmad M, Bennett IM, Azzoni L, Alnemri ES, Perussia B. Natural killer (NK) cell-mediated cytotoxicity: differential use of TRAIL and Fas ligand by immature and mature primary human NK cells. J Exp Med 1998; 188(12): 2375-80.
[http://dx.doi.org/10.1084/jem.188.12.2375] [PMID: 9858524] ]. Cytokines released from activated NK cells play an important role in NK cell effector functions. One important cytokine produced from activated NK cells is TNF-α which induces tumor-cell apoptosis [3Cheng M, Chen Y, Xiao W, Sun R, Tian Z. NK cell-based immunotherapy for malignant diseases. Cell Mol Immunol 2013; 10(3): 230-52.
[http://dx.doi.org/10.1038/cmi.2013.10] [PMID: 23604045] ]. IFN-γ is another important cytokine produced by activated NK cells is, which exerts antitumor functions in various ways, including inhibition of the proliferation of tumour cells, restriction of tumor angiogenesis and stimulation of the adaptive immunity [13Street SE, Cretney E, Smyth MJ. Perforin and interferon-gamma activities independently control tumor initiation, growth, and metastasis. Blood 2001; 97(1): 192-7.
[http://dx.doi.org/10.1182/blood.V97.1.192] [PMID: 11133760] , 14Yang G, Xu Y, Chen X, Hu G. IFITM1 plays an essential role in the antiproliferative action of interferon-gamma. Oncogene 2007; 26(4): 594-603.
[http://dx.doi.org/10.1038/sj.onc.1209807] [PMID: 16847454] ]. By expressing FcγRIIIa (type III receptor For Constant (Fc) region of IgG; CD16), NK cells can kill tumour cells by Antibody-Dependent Cellular Cytotoxicity (ADCC) [3Cheng M, Chen Y, Xiao W, Sun R, Tian Z. NK cell-based immunotherapy for malignant diseases. Cell Mol Immunol 2013; 10(3): 230-52.
[http://dx.doi.org/10.1038/cmi.2013.10] [PMID: 23604045] ]. CD16 is a NK cell activating receptor that enables NK cells to recognize IgG-coated tumor cells, resulting in their killing through the release of their cytotoxic granules containing perforins and granzymes at the immunological synapse [15Becknell B, Caligiuri MA. Natural Killer Cells in Innate Immunity and Cancer. J Immunotherapy 2008; 31: 685-92.]. Cytokines such as IL-2, IL-12, IL-18, or IL-15, released from other cells of the immune system can further enhance NK cell effector functions.
NK cells express an orchestra of activating and inhibitory receptors that dictate the attack and killing of cancer cells or “stressed” self-cells, while protecting normal cells. These receptors can either stimulate NK cell effector functions (activating receptors) or dampen NK cell reactivity towards a target cell (inhibitory receptors). The balance between the signals conveyed by activating and inhibitory receptors determines whether NK cells are activated to kill a target cell or not [6Vivier E, Raulet DH, Moretta A, et al. Innate or adaptive immunity? The example of natural killer cells. Science 2011; 331(6013): 44-9.
[http://dx.doi.org/10.1126/science.1198687] [PMID: 21212348] , 16Vivier E, Ugolini S, Blaise D, Chabannon C, Brossay L. Targeting natural killer cells and natural killer T cells in cancer. Nat Rev Immunol 2012; 12(4): 239-52.
[http://dx.doi.org/10.1038/nri3174] [PMID: 22437937] , 17Koch J, Steinle A, Watzl C, Mandelboim O. Activating natural cytotoxicity receptors of natural killer cells in cancer and infection. Trends Immunol 2013; 34(4): 182-91.
[http://dx.doi.org/10.1016/j.it.2013.01.003] [PMID: 23414611] ].
The ligands for the Inhibitory receptors are MHC class I molecules [18Moretta A, Bottino C, Vitale M, et al. Receptors for HLA class-I molecules in human natural killer cells. Annu Rev Immunol 1996; 14: 619-48.
[http://dx.doi.org/10.1146/annurev.immunol.14.1.619] [PMID: 8717527] , 19Moretta L, Ciccone E, Moretta A, Höglund P, Ohlén C, Kärre K. Allorecognition by NK cells: Nonself or no self? Immunol Today 1992; 13(8): 300-6.
[http://dx.doi.org/10.1016/0167-5699(92)90042-6] [PMID: 1380815] ]. By binding to MHC class I molecules, inhibitory receptors enable NK cells to discriminate between self and altered-self cells. Thus NK cells are “educated” or “licensed” during their development to mature into a functional repertoire of NK cells that are adapted to the MHC class I molecules of the host [16Vivier E, Ugolini S, Blaise D, Chabannon C, Brossay L. Targeting natural killer cells and natural killer T cells in cancer. Nat Rev Immunol 2012; 12(4): 239-52.
[http://dx.doi.org/10.1038/nri3174] [PMID: 22437937] ]. NK cell effector functions are inhibited by the engagement of the inhibitory receptors to self MHC class I molecules, thus NK cells are “tolerant” to healthy host cells. The strength of the activating signals NK cells receive upon encountering healthy cells is diminished by the engagement of inhibitory receptors to MHC class I molecules [15Becknell B, Caligiuri MA. Natural Killer Cells in Innate Immunity and Cancer. J Immunotherapy 2008; 31: 685-92.]. Malignant transformation can be associated with decreased expression of MHC class I molecules. NK cells become activated by cells with reduced expression of MHC class I molecules. This is because those cells are no longer kept in check by the engagement of the inhibitory receptors to MHC class I molecules. This hypothesis is known as the “missing-self” stimulation of NK cell activation [20Ljunggren HG, Kärre K. In search of the ‘missing self’: MHC molecules and NK cell recognition. Immunol Today 1990; 11(7): 237-44.
[http://dx.doi.org/10.1016/0167-5699(90)90097-S] [PMID: 2201309] ]. Inhibitory receptors specific to MHC class I molecules include Killer cell Immunoglobulin-like Receptors (KIRs) and lectin-like CD94/NKG2A heterodimers. While KIRs bind the classical MHC-I molecules (HLA-A, HLA-B and HLA-C molecules), CD94/NKG2A binds the non-classical MHC-Ib molecules (HLA-E in humans) [21He Y, Tian Z. NK cell education via nonclassical MHC and non-MHC ligands. Cell Mol Immunol 2017; 14(4): 321-30.
[http://dx.doi.org/10.1038/cmi.2016.26] [PMID: 27264685] ]. CD94/NKG2A is important in educating NK cell tolerance to self and contributes to the inhibition of antitumor immunity mediated by NK cells [21He Y, Tian Z. NK cell education via nonclassical MHC and non-MHC ligands. Cell Mol Immunol 2017; 14(4): 321-30.
[http://dx.doi.org/10.1038/cmi.2016.26] [PMID: 27264685] ]. These inhibitory receptors mediate their inhibitory functions by signaling through intracytoplasmic Immunoreceptor Tyrosine-based Inhibition Motifs (ITIMs) [18Moretta A, Bottino C, Vitale M, et al. Receptors for HLA class-I molecules in human natural killer cells. Annu Rev Immunol 1996; 14: 619-48.
[http://dx.doi.org/10.1146/annurev.immunol.14.1.619] [PMID: 8717527] , 19Moretta L, Ciccone E, Moretta A, Höglund P, Ohlén C, Kärre K. Allorecognition by NK cells: Nonself or no self? Immunol Today 1992; 13(8): 300-6.
[http://dx.doi.org/10.1016/0167-5699(92)90042-6] [PMID: 1380815] ]. These are conserved signaling domains that consist of six-amino acid sequences present in the cytoplasmic tails of the inhibitory receptors. Upon engagement of ITIM-containing inhibitory receptor to its specific MHC class I molecule, the tyrosine residue if the ITIM is phosphorylated by Src family kinases, resulting in the recruitment of the tyrosine phosphatases SHP-1, SHP-2, and SHIP-1, which in turn dephosphorylate the protein substrates of the tyrosine kinases linked to activating NK receptors. Consequently, degranulation, cytokine release and proliferation of NK cells are inhibited [22Lanier LL. Up on the tightrope: Natural killer cell activation and inhibition. Nat Immunol 2008; 9(5): 495-502.
[http://dx.doi.org/10.1038/ni1581] [PMID: 18425106] ].
In addition to the MHC-I dependent inhibitory receptors, KIRs and CD94/NKG2A, NK cells express additional inhibitory receptors that do not bind MHC-I molecules. These inhibitory receptors might also be involved in the NK cell education process. Among these inhibitory receptors, T cell immunoglobulin and ITIM domain (TIGIT) is expressed on NK cells as well as T cells. The ligand for TIGIT is CD155/PVR (the poliovirus receptor). Mouse TIGIT inhibits NK cell cytotoxicity upon interaction with PVR [23Stanietsky N, Rovis TL, Glasner A, et al. Mouse TIGIT inhibits NK-cell cytotoxicity upon interaction with PVR. Eur J Immunol 2013; 43(8): 2138-50.
[http://dx.doi.org/10.1002/eji.201243072] [PMID: 23677581] ]. Upregulation of TIGIT during liver regeneration, inhibited NK cell overactivation by interaction with CD155. This suggests that TIGIT plays a role in NK cell self-tolerance towards regenerative hyperplasia [23Stanietsky N, Rovis TL, Glasner A, et al. Mouse TIGIT inhibits NK-cell cytotoxicity upon interaction with PVR. Eur J Immunol 2013; 43(8): 2138-50.
[http://dx.doi.org/10.1002/eji.201243072] [PMID: 23677581] ]. TIGIT is significantly upregulated in tumor infiltrating T cells, marking them as chronically stimulated or exhausted tumor infiltrating T cells. As CD155 is highly expressed in several types of human malignancies [24Gao J, Zheng Q, Xin N, Wang W, Zhao C. CD155, an onco-immunologic molecule in human tumors. Cancer Sci 2017; 108(10): 1934-8.
[http://dx.doi.org/10.1111/cas.13324] [PMID: 28730595] ], NK cells might become similarly “tumor-tolerant” and become hyporesponsive by interaction with CD155. Another inhibitory receptor that can interact with CD155 is CD96 [25Chan CJ, Martinet L, Gilfillan S, et al. The receptors CD96 and CD226 oppose each other in the regulation of natural killer cell functions. Nat Immunol 2014; 15(5): 431-8.
[http://dx.doi.org/10.1038/ni.2850] [PMID: 24658051] ] CD96 is a co-inhibitory receptor that is expressed on NK as well as T cells [25Chan CJ, Martinet L, Gilfillan S, et al. The receptors CD96 and CD226 oppose each other in the regulation of natural killer cell functions. Nat Immunol 2014; 15(5): 431-8.
[http://dx.doi.org/10.1038/ni.2850] [PMID: 24658051] ].
NK cells also express surface activating receptors that detect internal changes that occur in damaged/stressed host tissues. This hypothesis is known as “stress-induced self” stimulation of NK cell activation [17Koch J, Steinle A, Watzl C, Mandelboim O. Activating natural cytotoxicity receptors of natural killer cells in cancer and infection. Trends Immunol 2013; 34(4): 182-91.
[http://dx.doi.org/10.1016/j.it.2013.01.003] [PMID: 23414611] , 26Kruse PH, Matta J, Ugolini S, Vivier E. Natural cytotoxicity receptors and their ligands. Immunol Cell Biol 2014; 92(3): 221-9.
[http://dx.doi.org/10.1038/icb.2013.98] [PMID: 24366519] ]. This mode of recognition relies on the detection of self molecules (ligands for the activating receptors) that are barely detectable in healthy cells but can be up-regulated by various forms of stress such as malignant transformation [16Vivier E, Ugolini S, Blaise D, Chabannon C, Brossay L. Targeting natural killer cells and natural killer T cells in cancer. Nat Rev Immunol 2012; 12(4): 239-52.
[http://dx.doi.org/10.1038/nri3174] [PMID: 22437937] ]. Activating receptors include CD16, C-type lectin-like receptors; NKG2D, NKp80, and the heterodimer CD94/NKG2C, the nectin binding adhesion molecule, DNAX Accessory Molecule 1 (DNAM1/CD226) activating Killer cell immunoglobulinlike Receptors (KAR), SLAM-Related Receptors (SRRs), NTB-A, CRACC and 2B4, and natural cytotoxicity receptors (NCRs); NKp30, NKp44, and NKp46 [27Joyce MG, Sun PD. The structural basis of ligand recognition by natural killer cell receptors. J Biomed Biotechnol 2011; 2011: 203628.
[http://dx.doi.org/10.1155/2011/203628] [PMID: 21629745] ]. Several activating receptors signal via adaptor proteins which contain an Immunoreceptor Tyrosine-based Activation Motif (ITAM). Engagement of activating receptors to their specific ligand on tumor cells leads to phosphorylation of the tyrosine residue of the ITAM resulting in recruitment of the tyrosine kinases Syk and ZAP70 which then trigger downstream events eventually leading to NK cell activation [22Lanier LL. Up on the tightrope: Natural killer cell activation and inhibition. Nat Immunol 2008; 9(5): 495-502.
[http://dx.doi.org/10.1038/ni1581] [PMID: 18425106] ].
3. THE FAMILY OF NATURAL CYTOTOXICITY RECEPTORS
The family of NCRs constitutes three type I membrane proteins that belong to the immunoglobulin superfamily [28Cantoni C, Bottino C, Vitale M, et al. NKp44, a triggering receptor involved in tumor cell lysis by activated human natural killer cells, is a novel member of the immunoglobulin superfamily. J Exp Med 1999; 189(5): 787-96.
[http://dx.doi.org/10.1084/jem.189.5.787] [PMID: 10049942] -31Pazina T, Shemesh A, Brusilovsky M, Porgador A, Campbell KS. Regulation of the Functions of Natural Cytotoxicity Receptors by Interactions with Diverse Ligands and Alterations in Splice Variant Expression. Front Immunol 2017; 8: 369.
[http://dx.doi.org/10.3389/fimmu.2017.00369] [PMID: 28424697] ]. The three NCRs: NKp46 (NCR1; CD335) [30Pessino A, Sivori S, Bottino C, et al. Molecular cloning of NKp46: A novel member of the immunoglobulin superfamily involved in triggering of natural cytotoxicity. J Exp Med 1998; 188(5): 953-60.
[http://dx.doi.org/10.1084/jem.188.5.953] [PMID: 9730896] ], NKp44 (NCR2; CD336) [28Cantoni C, Bottino C, Vitale M, et al. NKp44, a triggering receptor involved in tumor cell lysis by activated human natural killer cells, is a novel member of the immunoglobulin superfamily. J Exp Med 1999; 189(5): 787-96.
[http://dx.doi.org/10.1084/jem.189.5.787] [PMID: 10049942] ], and NKp30 (NCR3; CD337) [29Pende D, Parolini S, Pessino A, et al. Identification and molecular characterization of NKp30, a novel triggering receptor involved in natural cytotoxicity mediated by human natural killer cells. J Exp Med 1999; 190(10): 1505-16.
[http://dx.doi.org/10.1084/jem.190.10.1505] [PMID: 10562324] ] were discovered in the late 1990s as being expressed on human NK cells. All of the NCRs comprise one or two extracellular immunoglobulin-like ligand-binding domains, which binds to cellular and exogenously derived ligands, a transmembrane domain, and a short cytoplasmic domain [17Koch J, Steinle A, Watzl C, Mandelboim O. Activating natural cytotoxicity receptors of natural killer cells in cancer and infection. Trends Immunol 2013; 34(4): 182-91.
[http://dx.doi.org/10.1016/j.it.2013.01.003] [PMID: 23414611] , 26Kruse PH, Matta J, Ugolini S, Vivier E. Natural cytotoxicity receptors and their ligands. Immunol Cell Biol 2014; 92(3): 221-9.
[http://dx.doi.org/10.1038/icb.2013.98] [PMID: 24366519] ]. The NCRs lack an intracellular signaling domain and therefore associate with functional adaptor proteins containing ITAMs via a charged residue in their transmembrane domain [17Koch J, Steinle A, Watzl C, Mandelboim O. Activating natural cytotoxicity receptors of natural killer cells in cancer and infection. Trends Immunol 2013; 34(4): 182-91.
[http://dx.doi.org/10.1016/j.it.2013.01.003] [PMID: 23414611] , 26Kruse PH, Matta J, Ugolini S, Vivier E. Natural cytotoxicity receptors and their ligands. Immunol Cell Biol 2014; 92(3): 221-9.
[http://dx.doi.org/10.1038/icb.2013.98] [PMID: 24366519] ]. Unlike T Cell Receptors (TCRs) and immunoglobulins, NCRs do not undergo any somatic recombination [i.e., V(D) J genetic rearrangements] in order to become functionally active [32Hudspeth K, Silva-Santos B, Mavilio D. Natural cytotoxicity receptors: Broader expression patterns and functions in innate and adaptive immune cells. Front Immunol 2013; 4: 69.
[http://dx.doi.org/10.3389/fimmu.2013.00069] [PMID: 23518691] ].
3.1. NKp30
NKp30 is a 30 kDa protein that is expressed on all mature resting and activated human NK cells [29Pende D, Parolini S, Pessino A, et al. Identification and molecular characterization of NKp30, a novel triggering receptor involved in natural cytotoxicity mediated by human natural killer cells. J Exp Med 1999; 190(10): 1505-16.
[http://dx.doi.org/10.1084/jem.190.10.1505] [PMID: 10562324] ]. The gene encoding NKp30 is located on chromosome 6, in the class III region of the human MHC locus [29Pende D, Parolini S, Pessino A, et al. Identification and molecular characterization of NKp30, a novel triggering receptor involved in natural cytotoxicity mediated by human natural killer cells. J Exp Med 1999; 190(10): 1505-16.
[http://dx.doi.org/10.1084/jem.190.10.1505] [PMID: 10562324] ]. The NKp30 molecule is composed of one extracellular immunoglobulin-like ligand-binding domain. The transmembrane domain contains a charged arginine and the cytoplasmic domain has no additional signaling domain [33Joyce MG, Tran P, Zhuravleva MA, Jaw J, Colonna M, Sun PD. Crystal structure of human natural cytotoxicity receptor NKp30 and identification of its ligand binding site. Proc Natl Acad Sci USA 2011; 108(15): 6223-8.
[http://dx.doi.org/10.1073/pnas.1100622108] [PMID: 21444796] , 34Li Y, Wang Q, Mariuzza RA. Structure of the human activating natural cytotoxicity receptor NKp30 bound to its tumor cell ligand B7-H6. J Exp Med 2011; 208(4): 703-14.
[http://dx.doi.org/10.1084/jem.20102548] [PMID: 21422170] ]. Therefore, NKp30 associates with ITAM-containing adaptor signaling proteins, such as disulfide-linked homodimers of CD3ζ or heterodimers of CD3ζ with the γ-chain of the high-affinity Fc receptor for IgE (FcRγ) [17Koch J, Steinle A, Watzl C, Mandelboim O. Activating natural cytotoxicity receptors of natural killer cells in cancer and infection. Trends Immunol 2013; 34(4): 182-91.
[http://dx.doi.org/10.1016/j.it.2013.01.003] [PMID: 23414611] ]. Expression of NKp30 was found to be reduced in acute myeloid leukemia in one study [35Fauriat C, Just-Landi S, Mallet F, et al. Deficient expression of NCR in NK cells from acute myeloid leukemia: Evolution during leukemia treatment and impact of leukemia cells in NCRdull phenotype induction. Blood 2007; 109(1): 323-30.
[http://dx.doi.org/10.1182/blood-2005-08-027979] [PMID: 16940427] ] while another study documented no significant difference in NKp30 expression in Acute Myeloid Leukemia (AML) compared to healthy subjects [36Stringaris K, Sekine T, Khoder A, et al. Leukemia-induced phenotypic and functional defects in natural killer cells predict failure to achieve remission in acute myeloid leukemia. Haematologica 2014; 99(5): 836-47.
[http://dx.doi.org/10.3324/haematol.2013.087536] [PMID: 24488563] ]. This discrepancy in expression data confirms the need of further studies on larger cohort of samples to further classify patients according to the level of expression of NKp30. In cervical cancer NKp30 expression was previously found to be significantly reduced compared to healthy donors [37Garcia-Iglesias T, Del Toro-Arreola A, Albarran-Somoza B, et al. Low NKp30, NKp46 and NKG2D expression and reduced cytotoxic activity on NK cells in cervical cancer and precursor lesions. BMC Cancer 2009; 9: 186.
[http://dx.doi.org/10.1186/1471-2407-9-186] [PMID: 19531227] ]. In breast cancer, compared to healthy donors, one study documented no significant difference in expression of NKp30 [38Rady M, Watzl C, Claus M, Khorshid O, Mahran L, Abou-Aisha K. Altered expression of miR-181a and miR-146a does not change the expression of surface NCRs in human NK cells https://www.nature.com/articles/srep41381#supplementary-information2017.
[http://dx.doi.org/10.1038/srep41381] ], while two studies documented significantly reduced NKp30 expression [39Mamessier E, Sylvain A, Thibult ML, et al. Human breast cancer cells enhance self tolerance by promoting evasion from NK cell antitumor immunity. J Clin Invest 2011; 121(9): 3609-22.
[http://dx.doi.org/10.1172/JCI45816] [PMID: 21841316] , 40Nieto-Velázquez NG, Torres-Ramos YD, Muñoz-Sánchez JL, et al. Altered Expression of Natural Cytotoxicity Receptors and NKG2D on Peripheral Blood NK Cell Subsets in Breast Cancer Patients. Transl Oncol 2016; 9(5): 384-91.
[http://dx.doi.org/10.1016/j.tranon.2016.07.003] [PMID: 27641642] ]. In all the studies the investigators relied on flow cytometric analysis for measurement of surface expression of NCRs. The discrepancy in expression data from different studies confirms the need of further studies analyzing the surface expression of NKp30 in larger cohort of samples. As high interpatient variability in expression data of NKp30 and NCRs in general is indeed expected, further classification of patients according to the level of expression of NCRs along with the assessment of NK cell functions will further unravel the way NCRs impact NK cell responsiveness to cancer cells.
3.2. NKp44
NKp44 is 44-kD surface receptor that shares high similarity to the family of Triggering Receptors Expressed on Myeloid cells (TREM) [17Koch J, Steinle A, Watzl C, Mandelboim O. Activating natural cytotoxicity receptors of natural killer cells in cancer and infection. Trends Immunol 2013; 34(4): 182-91.
[http://dx.doi.org/10.1016/j.it.2013.01.003] [PMID: 23414611] , 26Kruse PH, Matta J, Ugolini S, Vivier E. Natural cytotoxicity receptors and their ligands. Immunol Cell Biol 2014; 92(3): 221-9.
[http://dx.doi.org/10.1038/icb.2013.98] [PMID: 24366519] ]. Similar to NKp30, the gene encoding NKp44 is located on chromosome 6, in the class III region of the human MHC locus [28Cantoni C, Bottino C, Vitale M, et al. NKp44, a triggering receptor involved in tumor cell lysis by activated human natural killer cells, is a novel member of the immunoglobulin superfamily. J Exp Med 1999; 189(5): 787-96.
[http://dx.doi.org/10.1084/jem.189.5.787] [PMID: 10049942] ]. NKp44 possesses one extracellular immunoglobulin-like ligand-binding domain that displays a typical V-type immunoglobulin domain, connected with a 64-amino acid-long stalk domain, a single transmembrane domain and a short cytoplasmic tail. NKp44 delivers its activating signals by coupling to a dimer of the ITAM-containing adaptor DNAX-activating protein 12 (DAP12) for downstream signal transduction [41Campbell KS, Yusa S, Kikuchi-Maki A, Catina TL. NKp44 triggers NK cell activaion through DAP12 association that is not influenced by a putative cytoplasmic inhibitory sequence. J Immunol (Baltimore, Md: 1950) 2004;: -906 2004; 172(5): 899-906.]. Interestingly however, the cytoplasmic tail contains an amino acid sequence (EILYHTVA), representing an ITIM (ITIM; V/IxYxxL/V) [42Cantoni C, Ponassi M, Biassoni R, et al. The three-dimensional structure of the human NK cell receptor NKp44, a triggering partner in natural cytotoxicity. Structure (London, England 1993) 2003; 11(5): 725-906.
[http://dx.doi.org/10.1016/S0969-2126(03)00095-9] ]. This suggests that NKp44 has the potential to deliver inhibitory signals as well. In fact, recent studies showed that the NKP44 ITIM can inhibit NK cell effector functions but depending on the ligand that binds to NKp44 [43Rosental B, Brusilovsky M, Hadad U, et al. Proliferating cell nuclear antigen is a novel inhibitory ligand for the natural cytotoxicity receptor NKp44. J Immunol (Baltimore, Md 1950) 2011; 187: 5702-906.
[http://dx.doi.org/10.4049/jimmunol.1102267] ]. This is in spite of the fact that initial reports showed that the NKp44 ITIM is not functional [41Campbell KS, Yusa S, Kikuchi-Maki A, Catina TL. NKp44 triggers NK cell activaion through DAP12 association that is not influenced by a putative cytoplasmic inhibitory sequence. J Immunol (Baltimore, Md: 1950) 2004;: -906 2004; 172(5): 899-906.]. NKp44 is expressed on activated NK cells only, contrary to NKp46 and NKp30 which are expressed on activated as well as resting NK cells [28Cantoni C, Bottino C, Vitale M, et al. NKp44, a triggering receptor involved in tumor cell lysis by activated human natural killer cells, is a novel member of the immunoglobulin superfamily. J Exp Med 1999; 189(5): 787-96.
[http://dx.doi.org/10.1084/jem.189.5.787] [PMID: 10049942] , 44Vitale M, Bottino C, Sivori S, et al. NKp44, a novel triggering surface molecule specifically expressed by activated natural killer cells, is involved in non-major histocompatibility complex-restricted tumor cell lysis. J Exp Med 1998; 187(12): 2065-72.
[http://dx.doi.org/10.1084/jem.187.12.2065] [PMID: 9625766] ]. In breast cancer two studies documented significantly increased expression of NKp44 compared to healthy controls [38Rady M, Watzl C, Claus M, Khorshid O, Mahran L, Abou-Aisha K. Altered expression of miR-181a and miR-146a does not change the expression of surface NCRs in human NK cells https://www.nature.com/articles/srep41381#supplementary-information2017.
[http://dx.doi.org/10.1038/srep41381] , 40Nieto-Velázquez NG, Torres-Ramos YD, Muñoz-Sánchez JL, et al. Altered Expression of Natural Cytotoxicity Receptors and NKG2D on Peripheral Blood NK Cell Subsets in Breast Cancer Patients. Transl Oncol 2016; 9(5): 384-91.
[http://dx.doi.org/10.1016/j.tranon.2016.07.003] [PMID: 27641642] ].
3.3. NKp46
NKp46, a 46-kD receptor, was the first NCR to be identified in NK cells and is so far the most specific marker reported for NK cells [17Koch J, Steinle A, Watzl C, Mandelboim O. Activating natural cytotoxicity receptors of natural killer cells in cancer and infection. Trends Immunol 2013; 34(4): 182-91.
[http://dx.doi.org/10.1016/j.it.2013.01.003] [PMID: 23414611] , 26Kruse PH, Matta J, Ugolini S, Vivier E. Natural cytotoxicity receptors and their ligands. Immunol Cell Biol 2014; 92(3): 221-9.
[http://dx.doi.org/10.1038/icb.2013.98] [PMID: 24366519] ]. The gene encoding NKp46 is located in the Leukocyte Receptor Complex (LRC) on human chromosome 19 (19q13.42) [30Pessino A, Sivori S, Bottino C, et al. Molecular cloning of NKp46: A novel member of the immunoglobulin superfamily involved in triggering of natural cytotoxicity. J Exp Med 1998; 188(5): 953-60.
[http://dx.doi.org/10.1084/jem.188.5.953] [PMID: 9730896] ]. The LRC includes as well the killer cell immunoglobulin-like receptor genes [26Kruse PH, Matta J, Ugolini S, Vivier E. Natural cytotoxicity receptors and their ligands. Immunol Cell Biol 2014; 92(3): 221-9.
[http://dx.doi.org/10.1038/icb.2013.98] [PMID: 24366519] ]. NKp46 possesses two C2-type immunoglobulin-like extracellular domains arranged at an 85˚ angle to each other and connected by a hinge region [45Foster CE, Colonna M, Sun PD. Crystal structure of the human natural killer (NK) cell activating receptor NKp46 reveals structural relationship to other leukocyte receptor complex immunoreceptors. J Biol Chem 2003; 278(46): 46081-6.
[http://dx.doi.org/10.1074/jbc.M308491200] [PMID: 12960161] , 46Ponassi M, Cantoni C, Biassoni R, et al. Structure of the human NK cell triggering receptor NKp46 ectodomain. Biochem Biophys Res Commun 2003; 309(2): 317-23.
[http://dx.doi.org/10.1016/j.bbrc.2003.08.007] [PMID: 12951052] ]. The cytoplasmic region constitutes a highly charged 25-amino-acid domain [26Kruse PH, Matta J, Ugolini S, Vivier E. Natural cytotoxicity receptors and their ligands. Immunol Cell Biol 2014; 92(3): 221-9.
[http://dx.doi.org/10.1038/icb.2013.98] [PMID: 24366519] ]. As all NCRs, the cytoplasmic domain lacks any signaling motif, however, activating signals are delivered by coupling of an ITAM-containing CD3ζ and FcRγ adaptor proteins to an arginine located in the transmembrane region of NKp46 [26Kruse PH, Matta J, Ugolini S, Vivier E. Natural cytotoxicity receptors and their ligands. Immunol Cell Biol 2014; 92(3): 221-9.
[http://dx.doi.org/10.1038/icb.2013.98] [PMID: 24366519] ]. NKp46 is expressed on resting as well as activated NK cells [30Pessino A, Sivori S, Bottino C, et al. Molecular cloning of NKp46: A novel member of the immunoglobulin superfamily involved in triggering of natural cytotoxicity. J Exp Med 1998; 188(5): 953-60.
[http://dx.doi.org/10.1084/jem.188.5.953] [PMID: 9730896] , 47Sivori S, Vitale M, Morelli L, et al. p46, a novel natural killer cell-specific surface molecule that mediates cell activation. J Exp Med 1997; 186(7): 1129-36.
[http://dx.doi.org/10.1084/jem.186.7.1129] [PMID: 9314561] ]. Expression of NKp46 was previously documented to be reduced in acute myeloid leukemia compared to healthy subjects [35Fauriat C, Just-Landi S, Mallet F, et al. Deficient expression of NCR in NK cells from acute myeloid leukemia: Evolution during leukemia treatment and impact of leukemia cells in NCRdull phenotype induction. Blood 2007; 109(1): 323-30.
[http://dx.doi.org/10.1182/blood-2005-08-027979] [PMID: 16940427] , 36Stringaris K, Sekine T, Khoder A, et al. Leukemia-induced phenotypic and functional defects in natural killer cells predict failure to achieve remission in acute myeloid leukemia. Haematologica 2014; 99(5): 836-47.
[http://dx.doi.org/10.3324/haematol.2013.087536] [PMID: 24488563] ] and cervical cancer [37Garcia-Iglesias T, Del Toro-Arreola A, Albarran-Somoza B, et al. Low NKp30, NKp46 and NKG2D expression and reduced cytotoxic activity on NK cells in cervical cancer and precursor lesions. BMC Cancer 2009; 9: 186.
[http://dx.doi.org/10.1186/1471-2407-9-186] [PMID: 19531227] ]. Three studies documented no significant difference in expression of NKp46 in breast cancer compared to healthy subjects [38Rady M, Watzl C, Claus M, Khorshid O, Mahran L, Abou-Aisha K. Altered expression of miR-181a and miR-146a does not change the expression of surface NCRs in human NK cells https://www.nature.com/articles/srep41381#supplementary-information2017.
[http://dx.doi.org/10.1038/srep41381] -40Nieto-Velázquez NG, Torres-Ramos YD, Muñoz-Sánchez JL, et al. Altered Expression of Natural Cytotoxicity Receptors and NKG2D on Peripheral Blood NK Cell Subsets in Breast Cancer Patients. Transl Oncol 2016; 9(5): 384-91.
[http://dx.doi.org/10.1016/j.tranon.2016.07.003] [PMID: 27641642] ].
4. NCR SIGNALING AND SIGNAL INTEGRATION
NCR signaling pathways rely on coupling to one of the known ITAM-containing signaling adaptor proteins [48Vivier E, Nunès JA, Vély F. Natural killer cell signaling pathways. Science 2004; 306(5701): 1517-9.
[http://dx.doi.org/10.1126/science.1103478] [PMID: 15567854] , 49Watzl C, Long EO. Signal transduction during activation and inhibition of natural killer cells. Current protocols in immunology / edited by [et al.] Chapter 11, Unit 11 19B
[http://dx.doi.org/10.1002/0471142735.im1109bs90 (2010).] ]. These adaptor proteins include disulfide-linked homodimers or heterodimers of CD3ζ and/or FcRγ chains for NKp30 and NKp46, and disulfide-linked homodimer DAP12 for NKp44. Engagement of an NCR to its ligand results in activation of ITAM-dependent signaling molecules including the Protein Tyrosine Kinases (PTKs) of the SRC family, which phosphorylate the ITAMs of the NCR adaptor proteins. Phosphorylation of ITAMs results in binding and activation of Src-homology domain 2 (SH2) of PTKs of the SYK family; zeta chain-associated protein kinase 70 (ZAP70) and spleen tyrosine kinase (SYK) [17Koch J, Steinle A, Watzl C, Mandelboim O. Activating natural cytotoxicity receptors of natural killer cells in cancer and infection. Trends Immunol 2013; 34(4): 182-91.
[http://dx.doi.org/10.1016/j.it.2013.01.003] [PMID: 23414611] , 48Vivier E, Nunès JA, Vély F. Natural killer cell signaling pathways. Science 2004; 306(5701): 1517-9.
[http://dx.doi.org/10.1126/science.1103478] [PMID: 15567854] , 49Watzl C, Long EO. Signal transduction during activation and inhibition of natural killer cells. Current protocols in immunology / edited by [et al.] Chapter 11, Unit 11 19B
[http://dx.doi.org/10.1002/0471142735.im1109bs90 (2010).] ]. These tyrosine kinases phosphorylate transmembrane adaptor molecules such as linker for the activation of T cells (LAT) and Non-T cell Activation Linker (NTAL) leading to the phosphorylation, activation, and association, of several signaling complexes, including phosphatidylinositol 3-kinase (PI3K), phospholipase C-γ (PLC-γ1 and PLCγ2), VAV2 and VAV3 proteins [17Koch J, Steinle A, Watzl C, Mandelboim O. Activating natural cytotoxicity receptors of natural killer cells in cancer and infection. Trends Immunol 2013; 34(4): 182-91.
[http://dx.doi.org/10.1016/j.it.2013.01.003] [PMID: 23414611] , 48Vivier E, Nunès JA, Vély F. Natural killer cell signaling pathways. Science 2004; 306(5701): 1517-9.
[http://dx.doi.org/10.1126/science.1103478] [PMID: 15567854] , 49Watzl C, Long EO. Signal transduction during activation and inhibition of natural killer cells. Current protocols in immunology / edited by [et al.] Chapter 11, Unit 11 19B
[http://dx.doi.org/10.1002/0471142735.im1109bs90 (2010).] ]. Activation of these signaling complexes induces Ca2+ flux and cytoskeletal reorganisation, ultimately resulting in NK cell degranulation and release of cytotoxic granules containing perforins and granzymes as well as secretion of cytokines such as IFNγ and TNFα [17Koch J, Steinle A, Watzl C, Mandelboim O. Activating natural cytotoxicity receptors of natural killer cells in cancer and infection. Trends Immunol 2013; 34(4): 182-91.
[http://dx.doi.org/10.1016/j.it.2013.01.003] [PMID: 23414611] ].
5. TUMOR-ASSOCIATED LIGANDS FOR NCRS
5.1. HLA-B-Associated Transcript 3
HLA-B-associated transcript 3 (BAT3) is a nuclear protein, also known as BCL2-associated anthanogene 6 (BAG-6) is one of the first identified tumor cell ligands for NKp30 [50Pogge von Strandmann E, Simhadri VR, von Tresckow B, et al. Human leukocyte antigen-B-associated transcript 3 is released from tumor cells and engages the NKp30 receptor on natural killer cells. Immunity 2007; 27(6): 965-74.
[http://dx.doi.org/10.1016/j.immuni.2007.10.010] [PMID: 18055229] ]. The gene encoding BAT3 is located on chromosome 6 in the same locus, the class III region of the human MHC locus as NKp30. NKp30 seems to interact at the C-terminus of BAT3 [51Binici J, Hartmann J, Herrmann J, et al. A soluble fragment of the tumor antigen BCL2-associated athanogene 6 (BAG-6) is essential and sufficient for inhibition of NKp30-dependent cytotoxicity of natural killer cells. J Biochem 2013; 288(48): 34295-03.
[http://dx.doi.org/10.1074/jbc.M113.483602] ]. Reduced expression of BAT3 mRNA and protein levels in HeLa cells resulted in decreased NK cell-mediated killing. Moreover, overexpression of BAT3 in LS174T cells (colon carcinoma cell line) resulted in enhanced NK cell-mediated killing [50Pogge von Strandmann E, Simhadri VR, von Tresckow B, et al. Human leukocyte antigen-B-associated transcript 3 is released from tumor cells and engages the NKp30 receptor on natural killer cells. Immunity 2007; 27(6): 965-74.
[http://dx.doi.org/10.1016/j.immuni.2007.10.010] [PMID: 18055229] ]. Additionally, BAT3 released from tumor cells in exosomal vesicles triggered cytokine secretion of NK cells (TNF-α, IFN-γ) [50Pogge von Strandmann E, Simhadri VR, von Tresckow B, et al. Human leukocyte antigen-B-associated transcript 3 is released from tumor cells and engages the NKp30 receptor on natural killer cells. Immunity 2007; 27(6): 965-74.
[http://dx.doi.org/10.1016/j.immuni.2007.10.010] [PMID: 18055229] ]. The stimulatory ability of BAT3 has also been confirmed in a multiple myeloma xenograft in vivo model where injection of BAT3 blocking antibodies resulted in decreased tumor rejection when peripheral blood NK cells were injected into nude mice [50Pogge von Strandmann E, Simhadri VR, von Tresckow B, et al. Human leukocyte antigen-B-associated transcript 3 is released from tumor cells and engages the NKp30 receptor on natural killer cells. Immunity 2007; 27(6): 965-74.
[http://dx.doi.org/10.1016/j.immuni.2007.10.010] [PMID: 18055229] ]. Recently also melanoma cell lines stimulated with RIG-I triggering the extracellular release of BAT3/BAG6-containing exosomal vesicles enhanced the cytotoxic potential of NK cells [52Daßler-Plenker J, Reiners KS, van den Boorn JG, et al. RIG-I activation induces the release of extracellular vesicles with antitumor activity. OncoImmunology 2016; 5(10): e1219827.
[http://dx.doi.org/10.1080/2162402X.2016.1219827] [PMID: 27853642] ]. Contrary to that overexpression of soluble BAT3 in the plasma of Chronic Lymphocytic Leukaemia (CLL) patients was associated with impaired NK cell activity [53Reiners KS, Topolar D, Henke A, et al. Soluble ligands for NK cell receptors promote evasion of chronic lymphocytic leukemia cells from NK cell anti-tumor activity. Blood 2013; 121(18): 3658-65.
[http://dx.doi.org/10.1182/blood-2013-01-476606] [PMID: 23509156] ]. Indeed BAT3 was released from CLL cells and cell lines in soluble form suppressed NK cell cytotoxicity suggesting that the immune evasion of the CLL cells from NK cell attack is due to dysregulated balance of exosomal vs soluble BAT3.
5.2. B7-H6
B7-H6, a member of the B7 family H6 is another tumor cell ligand for NKp30 [54Brandt CS, Baratin M, Yi EC, et al. The B7 family member B7-H6 is a tumor cell ligand for the activating natural killer cell receptor NKp30 in humans. J Exp Med 2009; 206(7): 1495-503.
[http://dx.doi.org/10.1084/jem.20090681] [PMID: 19528259] ]. Like other members of the B7 family, B7-H6 is a cell surface transmembrane protein that possesses two extracellular immunoglobulin domains [26Kruse PH, Matta J, Ugolini S, Vivier E. Natural cytotoxicity receptors and their ligands. Immunol Cell Biol 2014; 92(3): 221-9.
[http://dx.doi.org/10.1038/icb.2013.98] [PMID: 24366519] ]. The crystal structure of the NKp30 bound to B7-H6 has been described [34Li Y, Wang Q, Mariuzza RA. Structure of the human activating natural cytotoxicity receptor NKp30 bound to its tumor cell ligand B7-H6. J Exp Med 2011; 208(4): 703-14.
[http://dx.doi.org/10.1084/jem.20102548] [PMID: 21422170] ]. B7-H6 interacts with its front β-sheet of the V-like domain with the front and back β-sheets of C-like domain of NKp30 [34Li Y, Wang Q, Mariuzza RA. Structure of the human activating natural cytotoxicity receptor NKp30 bound to its tumor cell ligand B7-H6. J Exp Med 2011; 208(4): 703-14.
[http://dx.doi.org/10.1084/jem.20102548] [PMID: 21422170] ]. B7–H6 surface expression is restricted to both primary tumors and tumor cell lines, while neither healthy nor stressed cells appear to express B7-H6 [31Pazina T, Shemesh A, Brusilovsky M, Porgador A, Campbell KS. Regulation of the Functions of Natural Cytotoxicity Receptors by Interactions with Diverse Ligands and Alterations in Splice Variant Expression. Front Immunol 2017; 8: 369.
[http://dx.doi.org/10.3389/fimmu.2017.00369] [PMID: 28424697] , 54Brandt CS, Baratin M, Yi EC, et al. The B7 family member B7-H6 is a tumor cell ligand for the activating natural killer cell receptor NKp30 in humans. J Exp Med 2009; 206(7): 1495-503.
[http://dx.doi.org/10.1084/jem.20090681] [PMID: 19528259] , 55Textor S, Bossler F, Henrich KO, et al. The proto-oncogene Myc drives expression of the NK cell-activating NKp30 ligand B7-H6 in tumor cells. OncoImmunology 2016; 5(7): e1116674.
[http://dx.doi.org/10.1080/2162402X.2015.1116674] [PMID: 27622013] ]. Expression of B7–H6 on the surface of tumor cells enhances NKp30-mediated killing by NK cells [32Hudspeth K, Silva-Santos B, Mavilio D. Natural cytotoxicity receptors: Broader expression patterns and functions in innate and adaptive immune cells. Front Immunol 2013; 4: 69.
[http://dx.doi.org/10.3389/fimmu.2013.00069] [PMID: 23518691] ]. Contrary to that metalloprotease-mediated tumor cell shedding of soluble B7-H6 lead to decreased surface expression of B7-H6 on tumor cells and reduced NKp30-mediated recognition by NK cells [56Schlecker E, Fiegler N, Arnold A, et al. Metalloprotease-mediated tumor cell shedding of B7-H6, the ligand of the natural killer cell-activating receptor NKp30. Cancer Res 2014; 74(13): 3429-40.
[http://dx.doi.org/10.1158/0008-5472.CAN-13-3017] [PMID: 24780758] ]. In fact increased levels of soluble B7-H6 was detects in sera of patients with melanoma suggesting that this could be a mechanism of immune evasion in which tumor cells escape NK-mediated recognition by metalloprotease-mediated shedding of soluble B7-H6 [56Schlecker E, Fiegler N, Arnold A, et al. Metalloprotease-mediated tumor cell shedding of B7-H6, the ligand of the natural killer cell-activating receptor NKp30. Cancer Res 2014; 74(13): 3429-40.
[http://dx.doi.org/10.1158/0008-5472.CAN-13-3017] [PMID: 24780758] ]. Also it was observed that NKp30 downregulation in patient with ovarian cancer is presumably due to expression of the soluble form of B7-H6 in the tumor microenvironment resulting in chronic engagement of NKp30 to the soluble B7-H6 further providing an explanation of evasion of NK cell mediated immunity by tumor cells [57Pesce S, Tabellini G, Cantoni C, et al. B7-H6-mediated downregulation of NKp30 in NK cells contributes to ovarian carcinoma immune escape. OncoImmunology 2015; 4(4): e1001224.
[http://dx.doi.org/10.1080/2162402X.2014.1001224] [PMID: 26137398] ].
5.3. Proliferating Cell Nuclear Antigen
Proliferating Cell Nuclear Antigen (PCNA) is the first identified tumor cell ligand for NKp44 [43Rosental B, Brusilovsky M, Hadad U, et al. Proliferating cell nuclear antigen is a novel inhibitory ligand for the natural cytotoxicity receptor NKp44. J Immunol (Baltimore, Md 1950) 2011; 187: 5702-906.
[http://dx.doi.org/10.4049/jimmunol.1102267] ]. Expression of PCNA is restricted to the nucleus and in spite of the fact that it is strongly associated with cancer, it was found to be a direct tumor cell ligand for NKp44 [26Kruse PH, Matta J, Ugolini S, Vivier E. Natural cytotoxicity receptors and their ligands. Immunol Cell Biol 2014; 92(3): 221-9.
[http://dx.doi.org/10.1038/icb.2013.98] [PMID: 24366519] ]. PCNA is recruited by tumor cells from the nucleus to the immunological synapses, between tumor cells and NK cells [43Rosental B, Brusilovsky M, Hadad U, et al. Proliferating cell nuclear antigen is a novel inhibitory ligand for the natural cytotoxicity receptor NKp44. J Immunol (Baltimore, Md 1950) 2011; 187: 5702-906.
[http://dx.doi.org/10.4049/jimmunol.1102267] ]. In fact, PCNA expression was initially shown to inhibit NK cell-mediated lysis of tumor target cells even before it was described to be a ligand for NKp44 [26Kruse PH, Matta J, Ugolini S, Vivier E. Natural cytotoxicity receptors and their ligands. Immunol Cell Biol 2014; 92(3): 221-9.
[http://dx.doi.org/10.1038/icb.2013.98] [PMID: 24366519] ]. Enforced overexpression of PCNA resulted in a decreased INF-γ secretion by IL-2-activated primary NK cells [43Rosental B, Brusilovsky M, Hadad U, et al. Proliferating cell nuclear antigen is a novel inhibitory ligand for the natural cytotoxicity receptor NKp44. J Immunol (Baltimore, Md 1950) 2011; 187: 5702-906.
[http://dx.doi.org/10.4049/jimmunol.1102267] ]. Taken together, NKp44 can be regarded as an inhibitory receptor delivering inhibitory signals to NK cells when bound to PCNA. Interestingly, transfecting NK92 cells with NKp44 with a non-functional ITIM motif showed no reduced killing of target cells expressing PCNA in comparison to cells transfected with the wild-type NKp44 [43Rosental B, Brusilovsky M, Hadad U, et al. Proliferating cell nuclear antigen is a novel inhibitory ligand for the natural cytotoxicity receptor NKp44. J Immunol (Baltimore, Md 1950) 2011; 187: 5702-906.
[http://dx.doi.org/10.4049/jimmunol.1102267] ]. This suggests that reduced NK cell-mediated killing after the interaction between NKp44 and PCNA is mediated by the ITIM motif of NKp44 conveying inhibitory signals to NK cell [41Campbell KS, Yusa S, Kikuchi-Maki A, Catina TL. NKp44 triggers NK cell activaion through DAP12 association that is not influenced by a putative cytoplasmic inhibitory sequence. J Immunol (Baltimore, Md: 1950) 2004;: -906 2004; 172(5): 899-906.].
5.4. NKp44L
NKp44L is a recently identified activating tumor cell ligand for NKp44. The sequence of the new ligand partially resembles the sequence of the Mixed-Lineage Leukaemia protein 5 (MLL5) gene but lacks the last 5 exons of MLL5 [58Baychelier F, Sennepin A, Ermonval M, Dorgham K, Debré P, Vieillard V. Identification of a cellular ligand for the natural cytotoxicity receptor NKp44. Blood 2013; 122(17): 2935-42.
[http://dx.doi.org/10.1182/blood-2013-03-489054] [PMID: 23958951] ]. Instead, it contains a unique exon coding for the C-terminal region of NKp44L, which is important for the interaction between NKp44L and NKp44. Expression of NKp44L was not detected in the nucleus, contrary to MLL5 [26Kruse PH, Matta J, Ugolini S, Vivier E. Natural cytotoxicity receptors and their ligands. Immunol Cell Biol 2014; 92(3): 221-9.
[http://dx.doi.org/10.1038/icb.2013.98] [PMID: 24366519] ], but instead, was found to be restricted to the cell cytoplasm [58Baychelier F, Sennepin A, Ermonval M, Dorgham K, Debré P, Vieillard V. Identification of a cellular ligand for the natural cytotoxicity receptor NKp44. Blood 2013; 122(17): 2935-42.
[http://dx.doi.org/10.1182/blood-2013-03-489054] [PMID: 23958951] ]. Interestingly, the tissue distribution of MLL5 and NKp44L is also different. NKP44L expression was not detected in healthy tissues except lung and testis that showed marginal expression of NKp44L. However, expression of NKp44L was highly detected in several tumour cell lines [58Baychelier F, Sennepin A, Ermonval M, Dorgham K, Debré P, Vieillard V. Identification of a cellular ligand for the natural cytotoxicity receptor NKp44. Blood 2013; 122(17): 2935-42.
[http://dx.doi.org/10.1182/blood-2013-03-489054] [PMID: 23958951] ]. The expression data for NKP44L in solid and hematological malignancies are still lacking.
5.5. Heparan Sulphates
Different Heparan Sulphate (HS) sequences have been reported to bind to the three NCRs [17Koch J, Steinle A, Watzl C, Mandelboim O. Activating natural cytotoxicity receptors of natural killer cells in cancer and infection. Trends Immunol 2013; 34(4): 182-91.
[http://dx.doi.org/10.1016/j.it.2013.01.003] [PMID: 23414611] , 26Kruse PH, Matta J, Ugolini S, Vivier E. Natural cytotoxicity receptors and their ligands. Immunol Cell Biol 2014; 92(3): 221-9.
[http://dx.doi.org/10.1038/icb.2013.98] [PMID: 24366519] , 32Hudspeth K, Silva-Santos B, Mavilio D. Natural cytotoxicity receptors: Broader expression patterns and functions in innate and adaptive immune cells. Front Immunol 2013; 4: 69.
[http://dx.doi.org/10.3389/fimmu.2013.00069] [PMID: 23518691] ]. HS proteoglycans are expressed on all animal cells, but at different levels in normal tissues compared to tumour cells [26Kruse PH, Matta J, Ugolini S, Vivier E. Natural cytotoxicity receptors and their ligands. Immunol Cell Biol 2014; 92(3): 221-9.
[http://dx.doi.org/10.1038/icb.2013.98] [PMID: 24366519] ]. The binding pattern of NKp30 and NKp46 to HS is similar but their binding intensities vary a lot. Howeve, NKp44 showed a completely different binding pattern compared to NKp30 and NKp46. Generally, the three NCRs bind to HS proteoglycans containing two to three sulphate groups per disaccharide unit [26Kruse PH, Matta J, Ugolini S, Vivier E. Natural cytotoxicity receptors and their ligands. Immunol Cell Biol 2014; 92(3): 221-9.
[http://dx.doi.org/10.1038/icb.2013.98] [PMID: 24366519] ]. However, the level of HS surface expression on target cells doesn’t seem to influence the activation of NK cells. This suggests that HS doesn’t represent a primary ligand for NCRs [59Warren HS, Jones AL, Freeman C, Bettadapura J, Parish CR. Evidence that the cellular ligand for the human NK cell activation receptor NKp30 is not a heparan sulfate glycosaminoglycan. J Immunol (Baltimore, Md: 1950) 2005; 175: 207-175.
[http://dx.doi.org/10.1182/blood-2013-03-489054] ]. HS might, however act as coreceptors in complexes with other ligands to modulate NK cell activation [26Kruse PH, Matta J, Ugolini S, Vivier E. Natural cytotoxicity receptors and their ligands. Immunol Cell Biol 2014; 92(3): 221-9.
[http://dx.doi.org/10.1038/icb.2013.98] [PMID: 24366519] ] (Fig. 1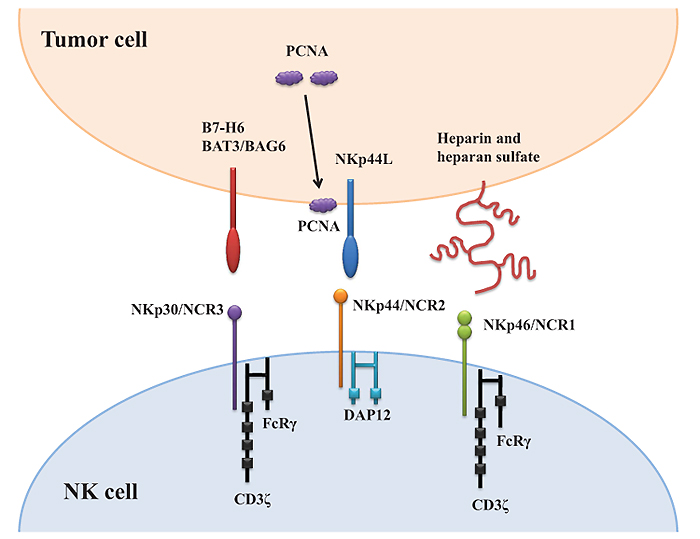 ).
).

CONCLUSIONS AND FUTURE PERSPECTIVES
Although NCRs were exclusively classified as NK cell activating receptors, several new findings show that interaction of NCRs with certain ligand can deliver inhibitory signals to NK cells or affect the recognition of tumor cells by NK cells. The fact that NCRs can interact with diverse ligands with little or no structural similarity implies that other ligands are yet to be explored. Moreover, clinical data about the expression of NCRs in NK cells along with their ligands whether on tumor cell surface or in the tumour microenvironment as soluble proteins and their impact on NK cell mediated antitumor immunity needs further studies. This will definitely aid in our understanding of novel mechanisms of the tumor escape from immune surveillance. Further studies manipulating NCRs’ expression in NK cells and tumor-associated ligands in tumor cells or the tumour microenvironment can further shape the strategies for the use of NK cells as an immunotherapeutic intervention for the treatment of cancer.
CONSENT FOR PUBLICATION
Not applicable.
CONFLICT OF INTEREST
The authors declare no conflict of interest, financial or otherwise.
ACKNOWLEDGEMENTS
Declared none.
REFERENCES
| [1] | Herberman RB, Nunn ME, Lavrin DH. Natural cytotoxic reactivity of mouse lymphoid cells against syngeneic acid allogeneic tumors. I. Distribution of reactivity and specificity. Int J Cancer 1975; 16(2): 216-29. [http://dx.doi.org/10.1002/ijc.2910160204] [PMID: 50294] |
| [2] | Kiessling R, Klein E, Pross H, Wigzell H. Natural killer cells in the mouse. II. Cytotoxic cells with specificity for mouse Moloney leukemia cells. Characteristics of the killer cell. Eur J Immunol 1975; 5(2): 117-21. [http://dx.doi.org/10.1002/eji.1830050209] [PMID: 1086218] |
| [3] | Cheng M, Chen Y, Xiao W, Sun R, Tian Z. NK cell-based immunotherapy for malignant diseases. Cell Mol Immunol 2013; 10(3): 230-52. [http://dx.doi.org/10.1038/cmi.2013.10] [PMID: 23604045] |
| [4] | Paust S, von Andrian UH. Natural killer cell memory. Nat Immunol 2011; 12(6): 500-8. [http://dx.doi.org/10.1038/ni.2032] [PMID: 21739673] |
| [5] | Sun JC, Lanier LL. NK cell development, homeostasis and function: Parallels with CD8+ T cells. Nat Rev Immunol 2011; 11(10): 645-57. [http://dx.doi.org/10.1038/nri3044] [PMID: 21869816] |
| [6] | Vivier E, Raulet DH, Moretta A, et al. Innate or adaptive immunity? The example of natural killer cells. Science 2011; 331(6013): 44-9. [http://dx.doi.org/10.1126/science.1198687] [PMID: 21212348] |
| [7] | Lanier LL, Testi R, Bindl J, Phillips JH. Identity of Leu-19 (CD56) leukocyte differentiation antigen and neural cell adhesion molecule. J Exp Med 1989; 169(6): 2233-8. [http://dx.doi.org/10.1084/jem.169.6.2233] [PMID: 2471777] |
| [8] | Ritz J, Schmidt RE, Michon J, Hercend T, Schlossman SF. Characterization of functional surface structures on human natural killer cells. Adv Immunol 1988; 42: 181-211. [http://dx.doi.org/10.1016/S0065-2776(08)60845-7] [PMID: 3284289] |
| [9] | Vivier E, Tomasello E, Baratin M, Walzer T, Ugolini S. Functions of natural killer cells. Nat Immunol 2008; 9(5): 503-10. [http://dx.doi.org/10.1038/ni1582] [PMID: 18425107] |
| [10] | Kägi D, Ledermann B, Bürki K, et al. Cytotoxicity mediated by T cells and natural killer cells is greatly impaired in perforin-deficient mice. Nature 1994; 369(6475): 31-7. [http://dx.doi.org/10.1038/369031a0] [PMID: 8164737] |
| [11] | Trapani JA, Davis J, Sutton VR, Smyth MJ. Proapoptotic functions of cytotoxic lymphocyte granule constituents in vitro and in vivo. Curr Opin Immunol 2000; 12(3): 323-9. [http://dx.doi.org/10.1016/S0952-7915(00)00094-7] [PMID: 10781403] |
| [12] | Zamai L, Ahmad M, Bennett IM, Azzoni L, Alnemri ES, Perussia B. Natural killer (NK) cell-mediated cytotoxicity: differential use of TRAIL and Fas ligand by immature and mature primary human NK cells. J Exp Med 1998; 188(12): 2375-80. [http://dx.doi.org/10.1084/jem.188.12.2375] [PMID: 9858524] |
| [13] | Street SE, Cretney E, Smyth MJ. Perforin and interferon-gamma activities independently control tumor initiation, growth, and metastasis. Blood 2001; 97(1): 192-7. [http://dx.doi.org/10.1182/blood.V97.1.192] [PMID: 11133760] |
| [14] | Yang G, Xu Y, Chen X, Hu G. IFITM1 plays an essential role in the antiproliferative action of interferon-gamma. Oncogene 2007; 26(4): 594-603. [http://dx.doi.org/10.1038/sj.onc.1209807] [PMID: 16847454] |
| [15] | Becknell B, Caligiuri MA. Natural Killer Cells in Innate Immunity and Cancer. J Immunotherapy 2008; 31: 685-92. |
| [16] | Vivier E, Ugolini S, Blaise D, Chabannon C, Brossay L. Targeting natural killer cells and natural killer T cells in cancer. Nat Rev Immunol 2012; 12(4): 239-52. [http://dx.doi.org/10.1038/nri3174] [PMID: 22437937] |
| [17] | Koch J, Steinle A, Watzl C, Mandelboim O. Activating natural cytotoxicity receptors of natural killer cells in cancer and infection. Trends Immunol 2013; 34(4): 182-91. [http://dx.doi.org/10.1016/j.it.2013.01.003] [PMID: 23414611] |
| [18] | Moretta A, Bottino C, Vitale M, et al. Receptors for HLA class-I molecules in human natural killer cells. Annu Rev Immunol 1996; 14: 619-48. [http://dx.doi.org/10.1146/annurev.immunol.14.1.619] [PMID: 8717527] |
| [19] | Moretta L, Ciccone E, Moretta A, Höglund P, Ohlén C, Kärre K. Allorecognition by NK cells: Nonself or no self? Immunol Today 1992; 13(8): 300-6. [http://dx.doi.org/10.1016/0167-5699(92)90042-6] [PMID: 1380815] |
| [20] | Ljunggren HG, Kärre K. In search of the ‘missing self’: MHC molecules and NK cell recognition. Immunol Today 1990; 11(7): 237-44. [http://dx.doi.org/10.1016/0167-5699(90)90097-S] [PMID: 2201309] |
| [21] | He Y, Tian Z. NK cell education via nonclassical MHC and non-MHC ligands. Cell Mol Immunol 2017; 14(4): 321-30. [http://dx.doi.org/10.1038/cmi.2016.26] [PMID: 27264685] |
| [22] | Lanier LL. Up on the tightrope: Natural killer cell activation and inhibition. Nat Immunol 2008; 9(5): 495-502. [http://dx.doi.org/10.1038/ni1581] [PMID: 18425106] |
| [23] | Stanietsky N, Rovis TL, Glasner A, et al. Mouse TIGIT inhibits NK-cell cytotoxicity upon interaction with PVR. Eur J Immunol 2013; 43(8): 2138-50. [http://dx.doi.org/10.1002/eji.201243072] [PMID: 23677581] |
| [24] | Gao J, Zheng Q, Xin N, Wang W, Zhao C. CD155, an onco-immunologic molecule in human tumors. Cancer Sci 2017; 108(10): 1934-8. [http://dx.doi.org/10.1111/cas.13324] [PMID: 28730595] |
| [25] | Chan CJ, Martinet L, Gilfillan S, et al. The receptors CD96 and CD226 oppose each other in the regulation of natural killer cell functions. Nat Immunol 2014; 15(5): 431-8. [http://dx.doi.org/10.1038/ni.2850] [PMID: 24658051] |
| [26] | Kruse PH, Matta J, Ugolini S, Vivier E. Natural cytotoxicity receptors and their ligands. Immunol Cell Biol 2014; 92(3): 221-9. [http://dx.doi.org/10.1038/icb.2013.98] [PMID: 24366519] |
| [27] | Joyce MG, Sun PD. The structural basis of ligand recognition by natural killer cell receptors. J Biomed Biotechnol 2011; 2011: 203628. [http://dx.doi.org/10.1155/2011/203628] [PMID: 21629745] |
| [28] | Cantoni C, Bottino C, Vitale M, et al. NKp44, a triggering receptor involved in tumor cell lysis by activated human natural killer cells, is a novel member of the immunoglobulin superfamily. J Exp Med 1999; 189(5): 787-96. [http://dx.doi.org/10.1084/jem.189.5.787] [PMID: 10049942] |
| [29] | Pende D, Parolini S, Pessino A, et al. Identification and molecular characterization of NKp30, a novel triggering receptor involved in natural cytotoxicity mediated by human natural killer cells. J Exp Med 1999; 190(10): 1505-16. [http://dx.doi.org/10.1084/jem.190.10.1505] [PMID: 10562324] |
| [30] | Pessino A, Sivori S, Bottino C, et al. Molecular cloning of NKp46: A novel member of the immunoglobulin superfamily involved in triggering of natural cytotoxicity. J Exp Med 1998; 188(5): 953-60. [http://dx.doi.org/10.1084/jem.188.5.953] [PMID: 9730896] |
| [31] | Pazina T, Shemesh A, Brusilovsky M, Porgador A, Campbell KS. Regulation of the Functions of Natural Cytotoxicity Receptors by Interactions with Diverse Ligands and Alterations in Splice Variant Expression. Front Immunol 2017; 8: 369. [http://dx.doi.org/10.3389/fimmu.2017.00369] [PMID: 28424697] |
| [32] | Hudspeth K, Silva-Santos B, Mavilio D. Natural cytotoxicity receptors: Broader expression patterns and functions in innate and adaptive immune cells. Front Immunol 2013; 4: 69. [http://dx.doi.org/10.3389/fimmu.2013.00069] [PMID: 23518691] |
| [33] | Joyce MG, Tran P, Zhuravleva MA, Jaw J, Colonna M, Sun PD. Crystal structure of human natural cytotoxicity receptor NKp30 and identification of its ligand binding site. Proc Natl Acad Sci USA 2011; 108(15): 6223-8. [http://dx.doi.org/10.1073/pnas.1100622108] [PMID: 21444796] |
| [34] | Li Y, Wang Q, Mariuzza RA. Structure of the human activating natural cytotoxicity receptor NKp30 bound to its tumor cell ligand B7-H6. J Exp Med 2011; 208(4): 703-14. [http://dx.doi.org/10.1084/jem.20102548] [PMID: 21422170] |
| [35] | Fauriat C, Just-Landi S, Mallet F, et al. Deficient expression of NCR in NK cells from acute myeloid leukemia: Evolution during leukemia treatment and impact of leukemia cells in NCRdull phenotype induction. Blood 2007; 109(1): 323-30. [http://dx.doi.org/10.1182/blood-2005-08-027979] [PMID: 16940427] |
| [36] | Stringaris K, Sekine T, Khoder A, et al. Leukemia-induced phenotypic and functional defects in natural killer cells predict failure to achieve remission in acute myeloid leukemia. Haematologica 2014; 99(5): 836-47. [http://dx.doi.org/10.3324/haematol.2013.087536] [PMID: 24488563] |
| [37] | Garcia-Iglesias T, Del Toro-Arreola A, Albarran-Somoza B, et al. Low NKp30, NKp46 and NKG2D expression and reduced cytotoxic activity on NK cells in cervical cancer and precursor lesions. BMC Cancer 2009; 9: 186. [http://dx.doi.org/10.1186/1471-2407-9-186] [PMID: 19531227] |
| [38] | Rady M, Watzl C, Claus M, Khorshid O, Mahran L, Abou-Aisha K. Altered expression of miR-181a and miR-146a does not change the expression of surface NCRs in human NK cells https://www.nature.com/articles/srep41381#supplementary-information2017. [http://dx.doi.org/10.1038/srep41381] |
| [39] | Mamessier E, Sylvain A, Thibult ML, et al. Human breast cancer cells enhance self tolerance by promoting evasion from NK cell antitumor immunity. J Clin Invest 2011; 121(9): 3609-22. [http://dx.doi.org/10.1172/JCI45816] [PMID: 21841316] |
| [40] | Nieto-Velázquez NG, Torres-Ramos YD, Muñoz-Sánchez JL, et al. Altered Expression of Natural Cytotoxicity Receptors and NKG2D on Peripheral Blood NK Cell Subsets in Breast Cancer Patients. Transl Oncol 2016; 9(5): 384-91. [http://dx.doi.org/10.1016/j.tranon.2016.07.003] [PMID: 27641642] |
| [41] | Campbell KS, Yusa S, Kikuchi-Maki A, Catina TL. NKp44 triggers NK cell activaion through DAP12 association that is not influenced by a putative cytoplasmic inhibitory sequence. J Immunol (Baltimore, Md: 1950) 2004;: -906 2004; 172(5): 899-906. |
| [42] | Cantoni C, Ponassi M, Biassoni R, et al. The three-dimensional structure of the human NK cell receptor NKp44, a triggering partner in natural cytotoxicity. Structure (London, England 1993) 2003; 11(5): 725-906. [http://dx.doi.org/10.1016/S0969-2126(03)00095-9] |
| [43] | Rosental B, Brusilovsky M, Hadad U, et al. Proliferating cell nuclear antigen is a novel inhibitory ligand for the natural cytotoxicity receptor NKp44. J Immunol (Baltimore, Md 1950) 2011; 187: 5702-906. [http://dx.doi.org/10.4049/jimmunol.1102267] |
| [44] | Vitale M, Bottino C, Sivori S, et al. NKp44, a novel triggering surface molecule specifically expressed by activated natural killer cells, is involved in non-major histocompatibility complex-restricted tumor cell lysis. J Exp Med 1998; 187(12): 2065-72. [http://dx.doi.org/10.1084/jem.187.12.2065] [PMID: 9625766] |
| [45] | Foster CE, Colonna M, Sun PD. Crystal structure of the human natural killer (NK) cell activating receptor NKp46 reveals structural relationship to other leukocyte receptor complex immunoreceptors. J Biol Chem 2003; 278(46): 46081-6. [http://dx.doi.org/10.1074/jbc.M308491200] [PMID: 12960161] |
| [46] | Ponassi M, Cantoni C, Biassoni R, et al. Structure of the human NK cell triggering receptor NKp46 ectodomain. Biochem Biophys Res Commun 2003; 309(2): 317-23. [http://dx.doi.org/10.1016/j.bbrc.2003.08.007] [PMID: 12951052] |
| [47] | Sivori S, Vitale M, Morelli L, et al. p46, a novel natural killer cell-specific surface molecule that mediates cell activation. J Exp Med 1997; 186(7): 1129-36. [http://dx.doi.org/10.1084/jem.186.7.1129] [PMID: 9314561] |
| [48] | Vivier E, Nunès JA, Vély F. Natural killer cell signaling pathways. Science 2004; 306(5701): 1517-9. [http://dx.doi.org/10.1126/science.1103478] [PMID: 15567854] |
| [49] | Watzl C, Long EO. Signal transduction during activation and inhibition of natural killer cells. Current protocols in immunology / edited by [et al.] Chapter 11, Unit 11 19B [http://dx.doi.org/10.1002/0471142735.im1109bs90 (2010).] |
| [50] | Pogge von Strandmann E, Simhadri VR, von Tresckow B, et al. Human leukocyte antigen-B-associated transcript 3 is released from tumor cells and engages the NKp30 receptor on natural killer cells. Immunity 2007; 27(6): 965-74. [http://dx.doi.org/10.1016/j.immuni.2007.10.010] [PMID: 18055229] |
| [51] | Binici J, Hartmann J, Herrmann J, et al. A soluble fragment of the tumor antigen BCL2-associated athanogene 6 (BAG-6) is essential and sufficient for inhibition of NKp30-dependent cytotoxicity of natural killer cells. J Biochem 2013; 288(48): 34295-03. [http://dx.doi.org/10.1074/jbc.M113.483602] |
| [52] | Daßler-Plenker J, Reiners KS, van den Boorn JG, et al. RIG-I activation induces the release of extracellular vesicles with antitumor activity. OncoImmunology 2016; 5(10): e1219827. [http://dx.doi.org/10.1080/2162402X.2016.1219827] [PMID: 27853642] |
| [53] | Reiners KS, Topolar D, Henke A, et al. Soluble ligands for NK cell receptors promote evasion of chronic lymphocytic leukemia cells from NK cell anti-tumor activity. Blood 2013; 121(18): 3658-65. [http://dx.doi.org/10.1182/blood-2013-01-476606] [PMID: 23509156] |
| [54] | Brandt CS, Baratin M, Yi EC, et al. The B7 family member B7-H6 is a tumor cell ligand for the activating natural killer cell receptor NKp30 in humans. J Exp Med 2009; 206(7): 1495-503. [http://dx.doi.org/10.1084/jem.20090681] [PMID: 19528259] |
| [55] | Textor S, Bossler F, Henrich KO, et al. The proto-oncogene Myc drives expression of the NK cell-activating NKp30 ligand B7-H6 in tumor cells. OncoImmunology 2016; 5(7): e1116674. [http://dx.doi.org/10.1080/2162402X.2015.1116674] [PMID: 27622013] |
| [56] | Schlecker E, Fiegler N, Arnold A, et al. Metalloprotease-mediated tumor cell shedding of B7-H6, the ligand of the natural killer cell-activating receptor NKp30. Cancer Res 2014; 74(13): 3429-40. [http://dx.doi.org/10.1158/0008-5472.CAN-13-3017] [PMID: 24780758] |
| [57] | Pesce S, Tabellini G, Cantoni C, et al. B7-H6-mediated downregulation of NKp30 in NK cells contributes to ovarian carcinoma immune escape. OncoImmunology 2015; 4(4): e1001224. [http://dx.doi.org/10.1080/2162402X.2014.1001224] [PMID: 26137398] |
| [58] | Baychelier F, Sennepin A, Ermonval M, Dorgham K, Debré P, Vieillard V. Identification of a cellular ligand for the natural cytotoxicity receptor NKp44. Blood 2013; 122(17): 2935-42. [http://dx.doi.org/10.1182/blood-2013-03-489054] [PMID: 23958951] |
| [59] | Warren HS, Jones AL, Freeman C, Bettadapura J, Parish CR. Evidence that the cellular ligand for the human NK cell activation receptor NKp30 is not a heparan sulfate glycosaminoglycan. J Immunol (Baltimore, Md: 1950) 2005; 175: 207-175. [http://dx.doi.org/10.1182/blood-2013-03-489054] |