- Home
- About Journals
-
Information for Authors/ReviewersEditorial Policies
Publication Fee
Publication Cycle - Process Flowchart
Online Manuscript Submission and Tracking System
Publishing Ethics and Rectitude
Authorship
Author Benefits
Reviewer Guidelines
Guest Editor Guidelines
Peer Review Workflow
Quick Track Option
Copyediting Services
Bentham Open Membership
Bentham Open Advisory Board
Archiving Policies
Fabricating and Stating False Information
Post Publication Discussions and Corrections
Editorial Management
Advertise With Us
Funding Agencies
Rate List
Kudos
General FAQs
Special Fee Waivers and Discounts
- Contact
- Help
- About Us
- Search




The Open Ornithology Journal
(Discontinued)
ISSN: 1874-4532 ― Volume 13, 2020
Cretaceous Reverie: Review of Birds of Stone: Chinese Avian Fossils from the Age of Dinosaurs by Luis M. Chiappe and Meng Qingjin1
Alan Feduccia*
Abstract
Birds of Stone contains a portfolio of outstanding photographs of the spectacularly preserved Jehol bird fossils, from the Chinese Lower Cretaceous, and other pertinent vertebrate fossils of varying ages, along with comments on each fossil. The book nicely illustrates a range of species of the radiation of enantiornithines (opposite birds), the dominant Mesozoic landbirds, as well as the ornithuromorphs, the Mesozoic antecedents of the modern neornithine birds. Although the first section of the book is fairly straight forward, the second section, on bird origins and their early evolution is one-sided, presenting only the popular paleontological view and omits discussion of controversial subjects. Examples are the highly speculative presence of dinosaur protofeathers and improbable scenarios of flight origins. There are no citations of the numerous credible opposing views in the literature.
Article Information

Identifiers and Pagination:
Year: 2018Volume: 11
First Page: 27
Last Page: 33
Publisher Id: TOOENIJ-11-27
DOI: 10.2174/1874453201811010027
Article History:
Received Date: 04/09/2017Revision Received Date: 10/01/2018
Acceptance Date: 29/01/2018
Electronic publication date: 12/02/2018
Collection year: 2018
open-access license: This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International Public License (CC-BY 4.0), a copy of which is available at: (https://creativecommons.org/licenses/by/4.0/legalcode). This license permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
* Address correspondence to this author at the Department of Biology, University of North Carolina, Chapel Hill, North Carolina 27599-3280, United States of America; Tel: 919-942-3377; E-mail: feduccia@bio.unc.edu
| Open Peer Review Details | |||
|---|---|---|---|
| Manuscript submitted on 04-09-2017 |
Original Manuscript | Cretaceous Reverie: Review of Birds of Stone: Chinese Avian Fossils from the Age of Dinosaurs by Luis M. Chiappe and Meng Qingjin1 | |
1. INTRODUCTION
Birds of Stone [1Chiappe LM. Glorified Dinosaurs: The Origin and Early Evolution of Birds 2007.] is a well-produced volume that contains beautiful color photos largely of fossil birds of the famous Lower Cretaceous Jehol Biota of northeastern China. It is a coffee-table style updated and expanded edition of Chiappe’s Glorified Dinosaurs: The Origin and Early Evolution of Birds (2007) [2Chiappe LM, Meng Q. Birds of Stone, Baltimore, MD, Johns Hopkins University Press 2016; pp 294 ] with additional photographs. The text likewise is an updated version of Chiappe (2007), adhering to general themes accepted by the American Museum of Natural History and its associates, now considered codified by many paleontologists. Their consensus view is that birds are living dinosaurs, that dinosaurs were endothermic and developed varied primitive feathers prior to the evolution of flight, for insulation or display. Almost all of the sophisticated aerodynamic architecture of birds, including a flight hand, pennaceous remiges, and fused and lightened skeleton evolved as exaptations in a context other than flight in earth-bound theropods. Text citations are lacking, as are contrary views, but there is a bibliography at the end.
The quality of the photography is outstanding, although the choice of subjects is somewhat unbalanced. While the photography is good, however, lack of scale bars makes it difficult for readers to know if the size of each bird is that of a turkey or that of a sparrow. Although more than 30 species of enantiornithines are known, there is disproportionate photographic representation of them. For example, there are numerous photos labeled “enantiornithine indeterminate” and some 26 pages with photos of Confuciusornis, yet some critical fossils are slighted, including the so-called “feathered dinosaur” Sinosauropteryx (two small photos) and the oviraptorosaur Caudipteryx (no photos), considered to be a secondarily flightless bird by many ornithologists and paleontologists. These two genera, along with the microraptors and anchiornithids are central to current debate on bird origins and their correct identity has yet to be established.
1.1. Sinosauropteryx and Protofeathers
According to Chiappe and Meng, this small Early Cretaceous compsognathid dinosaur, closely allied with the famous Late Jurassic Solnhofen Compsognathus, exhibits protofeathers following the argument that birds are dinosaurs; but others, disagree. Sinosauropteryx shows little affinity with birds and its dorsal mid-line filaments have no demonstrable feather affinity [3Lingham-Soliar T. The evolution of the feather: Sinosauropteryx, a colourful tail. J Orn 2011; 152: 567-77.
[http://dx.doi.org/10.1007/s10336-010-0620-y] -6Lingham-Soliar T, Feduccia A, Wang X. A new Chinese specimen indicates that ‘protofeathers’ in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibers. Proc Royal Soc Long B. Biol Sci 2007; 274: 1823-9.
[http://dx.doi.org/10.1098/rspb.2007.0352] ]. Much of the controversy arises from the fact that there are a number of Early Cretaceous fossils, namely Caudipteryx and microraptors, now considered theropods, that exhibit true avian pennaceous flight feathers, some with symmetric and some with asymmetric vanes. Given the incredible aerodynamic fine structure of avian pennaceous feathers [7Lingham-Soliar T, Murugan N. A new helical crossed-fibre structure of β-Keratin in flight feathers and Its biomechanical implications. PLoS One 2013; 8(6): e65849.
[http://dx.doi.org/10.1371/journal.pone.0065849] ], it is almost inconceivable that such integumentary appendages could have evolved in a non-flight context, and exaptational explanations are sadly deficient. Many of the fossil specimens with true avian feathers are also thought to exhibit “protofeathers.” However, a superior explanation is that such putative protofeathers represent avian pennaceous feathers that have been structurally simplified through taphonomic deterioration, maceration or other diagenetic fossilization processes. Such simplified structures may come to resemble what some workers have identified as protofeathers; so great caution, sadly missing to date, must be exercised in such assessments. These filaments may be dermal collagen fibers, and it is clear that Sinosauropteryx was actually covered with small tuberculated dinosaurian scales [8Lingham-Soliar T. The evolution of the feather: Scales on the tail of Sinosauropteryx and an interpretation of the dinosaur’s opisthotonic posture. J Orn 2013; 154: 455-63.
[http://dx.doi.org/10.1007/s10336-012-0910-7] ].
Collagen is the most ubiquitous structural protein and the primary protein of the connective tissue in all animals, comprising some 25% to 35% of whole-body protein in vertebrates (fibrous tissues, including tendons, ligaments, cartilage and bone, as well as skin). Depending on the degree of mineralization collagen tissues may be rigid (bone) or compliant (tendon), and dermal collagen meshworks characterize the sauropsid skin. Collagen is of universal occurrence in animals and it has remarkable structural integrity. Decomposing collagen fiber bundles have been shown experimentally in the hypodermis of a dolphin (Tursiops) [4Lingham-Soliar T. The dinosaurian origin of feathers: Perspective from dolphin (Cetacea) collagen fibers. Naturwissenschaften 2003; 90: 563-7.
[http://dx.doi.org/10.1007/s00114-003-0483-7] ], (Fig. 1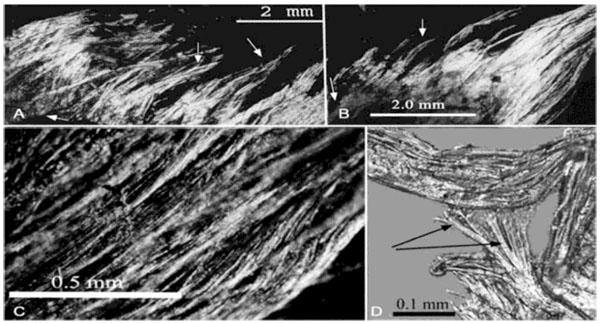 ), in ichthyosaurs [9Lingham-Soliar T. Rare soft tissue preservation showing fibrous structures in an ichthyosaur from the Lower Lias (Jurassic) of England. Proc Biol Sci 1999; 266: 2367-73.
), in ichthyosaurs [9Lingham-Soliar T. Rare soft tissue preservation showing fibrous structures in an ichthyosaur from the Lower Lias (Jurassic) of England. Proc Biol Sci 1999; 266: 2367-73.
[http://dx.doi.org/10.1098/rspb.1999.0933] ], and most recently in a Jurassic plesiosaur [10Vincent P, Allemand R, Taylor PD, Suan G, Maxwell EE. New insights on the systematics, palaeoecoloty and palaeobiology of a plesiosaurian with soft tissue preservation from the Toarcian of Holzmaden, Germany. Sci Nat 2017; 104: 51-00.
[http://dx.doi.org/10.1007/s00114-017-1472-6] ] and is of course the predominant structural protein in dinosaur skin. Collagen should always be considered the most likely protein when considering the identity of fibers in any vertebrate fossil preservation. To date there is no acceptable rigorous scientific evidence for the existence of protofeathers in dinosaurs or birds. Based on current evidence the best explanation for these varied filamentous structures is that they represent integumental meshworks of degraded collagen fibers, or in the case of animals with pennaceous avian feathers, the result of taphonomic alteration.
 |
Fig. (1) Putative but unproven “dinosaur protofeathers” strongly resemble dermal collagen fibers [6Lingham-Soliar T, Feduccia A, Wang X. A new Chinese specimen indicates that ‘protofeathers’ in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibers. Proc Royal Soc Long B. Biol Sci 2007; 274: 1823-9. [http://dx.doi.org/10.1098/rspb.2007.0352] ], shown here from (a) skin of a decomposing dophin [4Lingham-Soliar T. The dinosaurian origin of feathers: Perspective from dolphin (Cetacea) collagen fibers. Naturwissenschaften 2003; 90: 563-7. [http://dx.doi.org/10.1007/s00114-003-0483-7] ]. Protofeathers or collagen fibers? Decomposing collagen fiber bundles in the hypodermis of a dolphin, Tursiops. (A, B) Plume-like patterns of some of the fibers along the edges of the decomposing tissue. Many groups of fibers have narrow points of attachment (bottom arrow) that broaden before tapering again (due to degradation), giving the flame-like shape (top arrows). (C) Thick fiber bundles showing disorganization of the finer fibers and bundles to produce overlapping featherlike patterns. D) Detail of large fiber bundles showing component fibers during degradation, producing branching patterns among the filaments (adapted and modified from Lingham-Soliar4, courtesy Lingham-Soliar). |
1.2. Microraptor
Microraptor was the first four-wing “tetrapteryx” glider discovered from the Jehol biota. It was described as a small maniraptoran [11Xu X, Zhou Z, Wang X, Kuang X, Zhang F, Du X. Four-winged dinosaurs from China. Nature 2003; 421: 335-40.
[http://dx.doi.org/10.1038/nature01342] ], a theropod, and even as a basal dromaeosaurid. However, new evidence shows many avian characters, including an avian flight hand, a propatagium and asymmetric avian remiges [12Gong E-P, Martin LD, Burnham DA, Falk AF, Hou L-h. A new species of Microraptor from the jehol biota of northeastern china. Palaeoworld 2012; 21: 81-91.
[http://dx.doi.org/10.1016/j.palwor.2012.05.003] -15James F, Pourtless JA. Cladistics and the origin of birds: A review and two new analyses. Orn Monographs 2009; 66: 1-78.
[http://dx.doi.org/10.1525/om.2009.66.1.1] ]. Chiappe and Meng show a reconstruction of Microraptor in flight with the hind legs dropping below the body (p. 246), in keeping with an upright dinosaur posture, and they interpret hind-limb feathers as “trailing behind the lower leg and assisting in flight control (p.211).” This interpretation is in contrast the more logical explanation that the hind-limb asymmetric remiges formed a wing surface contributing to the overall planform. The hind-limb remiges are shown far too short; if accurate (p.211 photo), wingtips would drag on the ground if it ever ventured from trees, as many paleontologists believe. More probably, Microraptor was as originally described by Xu et al. [11Xu X, Zhou Z, Wang X, Kuang X, Zhang F, Du X. Four-winged dinosaurs from China. Nature 2003; 421: 335-40.
[http://dx.doi.org/10.1038/nature01342] ], a tetrapteryx glider with an aerodynamically refined sprawled, gliding posture. This interpretation is supported by the posture of most fossils (p. 247), casts of pelvic bones, and biomechanical comparisons to other vertebrate gliders, including mammals, and even human parachutists. Microraptor’s highly recurved manual claws with their needle-like tips (Fig. 2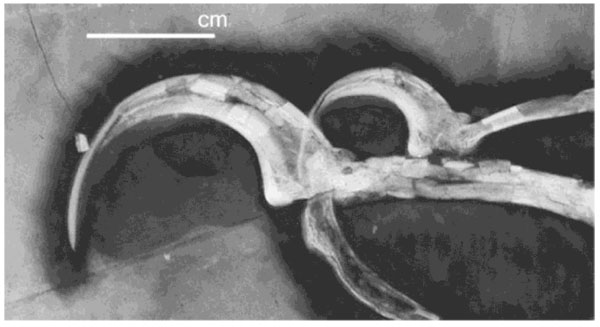 ) strongly suggest trunk-climbing habits, a fundamental avian adaptation [12Gong E-P, Martin LD, Burnham DA, Falk AF, Hou L-h. A new species of Microraptor from the jehol biota of northeastern china. Palaeoworld 2012; 21: 81-91.
) strongly suggest trunk-climbing habits, a fundamental avian adaptation [12Gong E-P, Martin LD, Burnham DA, Falk AF, Hou L-h. A new species of Microraptor from the jehol biota of northeastern china. Palaeoworld 2012; 21: 81-91.
[http://dx.doi.org/10.1016/j.palwor.2012.05.003] -14Burnham DA, Feduccia A, Martin LD, Falk AR. Tree climbing: A fundamental avian adaptation. J Syst Palaeontology 2011; 9: 103-7.
[http://dx.doi.org/10.1080/14772019.2010.522201] ] known for Archaeopteryx and many other basal birds. The view that “these animals would have dislocated their legs if they had adopted . . . a sprawled posture (p. 246)” is a stretch of biological credulity. Microraptor was most probably a bird, an offshoot of the early avian radiation of gliders, active fliers and flightless birds [14Burnham DA, Feduccia A, Martin LD, Falk AR. Tree climbing: A fundamental avian adaptation. J Syst Palaeontology 2011; 9: 103-7.
[http://dx.doi.org/10.1080/14772019.2010.522201] , 15James F, Pourtless JA. Cladistics and the origin of birds: A review and two new analyses. Orn Monographs 2009; 66: 1-78.
[http://dx.doi.org/10.1525/om.2009.66.1.1] ].
 |
Fig. (2) UV photograph of the manual claws of a Microraptor showing the keratinous sheath covering unguals III and IV [14Burnham DA, Feduccia A, Martin LD, Falk AR. Tree climbing: A fundamental avian adaptation. J Syst Palaeontology 2011; 9: 103-7. [http://dx.doi.org/10.1080/14772019.2010.522201] ]. These highly adapted and strongly curved and laterally compressed manual claws are typical of trunk-climbing early birds such as Archaeopteryx, as well as claws of woodpeckers and other scansorial birds, as well as mammals such as squirrels, etc., and provide conclusive evidence of trunk-climbing habits in this early avian “tetrapteryx” offshoot. (Courtesy D. Burnham). |
1.3. Caudipteryx
Chiappe and Meng consider Caudipteryx to have been a true theropod. However, numerous papers beginning in 2002 argue that it is avian, based on morphology and cladistic analyses, and important avian characters such as avian teeth and an avian skull with a postero-ventral foramen magnum, avian flight wings with a modern avian phalangeal formula, an avian arrangement of the remiges, and modern avian tail molt. Caudipteryx had scant “theropod” synapomorphies and like Archaeoperyx it lacked the definitive supra-acetabular crest, present in all true theropods. Some 20 publications beginning with Maryańska et al. [16Feduccia A. Fantasy vs Reality: A critique of Smith et al.’s Bird Origins. Open J Orn 2016; 9: 14-38.
[http://dx.doi.org/10.2174/1874453201609010014] -19Feduccia A. Riddle of the Feathered Dragons 2012.] show an avian status for Caudipteryx. Unfortunately, none are listed in the bibliography of Birds of Stone.
Conflation of birds and theropods, illustrated by Chiappe and others, has produced a hazardous arena for comparing everything from genome size to evolutionary trends. For Chiappe Caudipteryx is viewed as a flightless maniraptoran that developed advanced pennaceous avian remiges in a non-flight context, yet the arrangement of primaries and their attachment to manual skeletal elements is exactly like that of modern birds, and the phalangeal formula (2,3,2) is that of a modern bird. Its formerly volant ancestry is also suggested by the discovery of the more primitive, but closely allied Protarchaeopteryx. It had elongated forelimbs (wings) and was “probably predominately a ground-dwelling bird, but with some flight capability, like the living South American seriemas [19Feduccia A. Riddle of the Feathered Dragons 2012.].” Thus, Chiappe ignores the probable pedomorphic origin of Protarchaeopteryx and Caudipteryx and sees these taxa as learning to fly from the ground. Another problem attends the theropod Sinosauropteryx, with rudimentary forelimbs reduced to more than half the length of the hindlimbs. For either of these taxa to have anything to do with bird ancestry one must envision the re-elongation of already foreshortened forelimbs, a proposal completely at odds with Dollo’s Law. Since Chiappe denies any validity to Dollo’s Law (rule) [20Goldberg EE, Igi’c B. On phylogenetic tests of irreversible evolution. Evolution 2008; 62: 2727-41.
[http://dx.doi.org/10.1111/j.1558-5646.2008.00505.x] ], for Chiappe re-elongating forelimbs is no problem [21Chiappe LM. The dinosaur conspiracy. Bioscience 2012; 62(8): 770-2.
[http://dx.doi.org/10.1525/bio.2012.62.8.11] ]; yet, it remains a very thorny issue [19Feduccia A. Riddle of the Feathered Dragons 2012.]. The most reasonable explanation is that, contra Chiappe, Caudipteryx was a secondarily flightless bird that initially inherited its highly sophisticated "Flight characteristics from its volant ancestors". Oviraptorosaurs likely represent a Cretaceous radiation of ecologically, ratite-like secondarily flightless birds.
If Caudipteryx and other flightless taxa developed avian sophisticated aerodynamic architecture as exaptations, the model is basically non-Darwinian. The current exaptational explanations almost universally employed by Chiappe and other paleontologists “are often not fully formulated and rarely offer a biologically plausible hypothesis to account for their origin [15:31]”. “If most birdlike maniraptoran clades belong within Aves, problematic exaptational explanations including those for the origin of flight feathers, are unnecessary [15:31].” Given the aerodynamic perfection of the remarkably complex pennaceous remiges, it is inconceivable that such structures arose in a non-flight context.
1.4. Confuciusornis and Anchiornis
New studies of the earliest known beaked bird Confuciusornis (and Eoconfuciusornis) and the supposed troodontid theropod Anchiornis can be found in Falk et al., Zheng et al. [22Falk AR, Kaye TG, Zhou Z, Burnham DA. Laser fluorescence illuminates the soft tissue and life habits of the Early Cretaceous bird Confuciusornis. PLoS One 2016; 11(12): e0167284.
[http://dx.doi.org/10.1371/journal.pone.0167284] , 23Zheng X, O’Conner JK, Wang X, et al. Exceptional preservation of soft tissue in a new specimen of Eoconfuciusornis and its biological implications. Natl Sci Rev 2017; 4: 441-52. [doi: 10.1093/nst/nwx004].
[http://dx.doi.org/10.1093/nsr/nwx004] ] and Wang et al. [24Wang X, Pittman M, Zheng X, Kaye TG. Anchiornis soft tissues revealed by laser-stimulated fluorescence. Nat Commun 2017; 8: 14576a.
[http://dx.doi.org/10.1038/ncomms14576] ], illustrated by beautifully executed high-tech laser-stimulated fluorescence imaging. The former two studies leave no doubt that Confuciusornis was a fully volant arboreal perching bird, certainly not primarily terrestrial, as reconstructed by Chiappe [25Padian KL, Chiappe LM. The origin of birds and their flight. Sci Am 1998; 279: 38-47.
[http://dx.doi.org/10.1038/scientificamerican0298-38] , 26Chiappe LM, Ji S-A, Qiang J, Norell MA. Anatomy and systematics of the Confuciusornithidae (Theropoda, Aves) from the late Mesozoic of northeastern China. 1999.], and here pp. 8, 227, 230. As a note of caution it flew with “a primitive scapulocoracoid without an enlarged acrocoracoid process, it was flying without a keeled sternum, it was flying without an alula, and without a fully developed avian wrist” [27Olson SL. Review of anatomy and systematics of the confuciusornithidae (Theropoda: Aves) from the late mesozoic of China. Auk 2000; 117(3): 836-9.]. If Confuciusornis, a primitive bird quite basal on the avian tree, "possessed a suite of characters so modern, it suggests that these features arose much earlier than perhaps previously implied [22:1].” Anchiornis, considered to have been a troodontid dinosaur, is best interpreted as an archaeopterygid bird, as originally thought, closely allied with the urvogel Archaeopteryx. It was certainly a tetrapteryx glider and its soft anatomy was amazingly avian, including a pre and post-patagium, and avian arrangement of remiges [24Wang X, Pittman M, Zheng X, Kaye TG. Anchiornis soft tissues revealed by laser-stimulated fluorescence. Nat Commun 2017; 8: 14576a.
[http://dx.doi.org/10.1038/ncomms14576] ]. How could these avian structures have arisen in an earth-bound theropod? These recent papers about fine anatomy render much of the information about Confuciusornis and Anchiornis in Birds of Stone obsolete, and there is little reason to consider Anchiornis as anything other than a part of the early avian radiation.
2. FLIGHT ORIGINS
Questions concerning reconstructions and flight models for Chiappe and Meng derive strictly from the consensus view: Birds are living dinosaurs and therefore early basal birds are characterized as terrestrial obligate bipeds. Chiappe and Meng picture the secondarily flightless Caudipteryx, as “learning” to fly from the ground up, ignoring the fact that it is likely a flightless bird. They admit uncertainties concerning theories of flight origins, but advocate the improbable ground-up “gravity-resisted” origin of flight, following the “birds are dinosaurs” theme, but these models have been falsified time and time again [16Feduccia A. Fantasy vs Reality: A critique of Smith et al.’s Bird Origins. Open J Orn 2016; 9: 14-38.
[http://dx.doi.org/10.2174/1874453201609010014] -19Feduccia A. Riddle of the Feathered Dragons 2012., 28Evangelista D, Cam S, Huynh T, et al. Shifts in stability and control effectiveness during evolution of Paraves support aerial maneuvering hypotheses for flight origins. PeerJ 2014; 2: e632.
[http://dx.doi.org/10.7717/peerj.632] -30Alexander DE. On the Wing: Insects, pterosaurs, birds, bats and the evolution of animal flight 2015.]. They reject the most reasonable, “gravity-assisted” model, known as “wing-assisted climbing,” or WAC by Chatterjee, whose Johns Hopkins 2015 book [29Chatterjee S. The Rise of Birds 2nd ed. 2015.] is not listed in the bibliography. The ground-up flight scenarios favored by Chiappe are fatally flawed because the anatomy of early birds and their archosaurian or theropodan antecedents lack any significant pectoral apparatus and musculature for rigorous forelimb flapping to enact the proposed required movement. All these scenarios are not consistent with vertebrate flight origins in other groups, none of which developed flight from the ground up, only by the arboreal model, taking advantage of the cheap energy provided by gravity and high places. The arboreal theory is the most probable, “gravity-assisted” explanation, as opposed to the highly improbable “gravity-resisted” ground-up models, including Chiappe’s “ground-down” model. “A biomechanically parsimonious hypothesis for the evolution of flapping flight in terrestrial vertebrates suggests progression within an arboreal context from jumping to directed aerial descent, gliding with control via appendicular motions, and ultimately to powered flight [31Dudley R, Byrnes G, Yanoviak SP, Borrell B, Brown RM, McGuire JA. Gliding and the functional origins of flight: biomechanical novelty or necessity? Annu Rev Ecol Evol Syst 2007; 38: 179-201.
[http://dx.doi.org/10.1146/annurev.ecolsys.37.091305.110014] ].”
3. MINIATURIZATION AND DIGITAL FRAME SHIFT
Chiappe and Meng note that “The evolutionary transition from large dinosaurs to birds involved a dramatic miniaturization in which the forerunners of living birds were downsized as the result of a prolonged trend [emphasis mine] that started more than 50 million years prior to the appearance of the earliest birds (p. 73). This statement appears to resurrect the old orthogenesis, directional evolution toward some goal. Aside from obvious teleology, pedomorphosis is invoked to show that birds have juvenile dinosaur skulls with enlarged eyes and brains, but would better fit archosaurian ancestors. Heterochrony involving shorter developmental periods never changes only one aspect of morphology, and invariably results in much shorter forelimbs, not elongate wings thought to result from such a phenomenon. A pedomorphic event would have just the opposite effect as that desired for elongation of wings [19Feduccia A. Riddle of the Feathered Dragons 2012.]. Chiappe shows “compsognathids” (Sinosauropteryx) at the base of the avian tree, and, as noted, having abandoned Dollo’s Rule (Law) he apparently has no problem envisioning the improbable re-elongation of already foreshortened forelimbs.
Not discussed in Birds of Stone (but covered in Chiappe’s earlier book [2Chiappe LM, Meng Q. Birds of Stone, Baltimore, MD, Johns Hopkins University Press 2016; pp 294 ], but among the most controversial problems in the bird–dino debate has been the fact that birds embryologically have manual digits II-III-IV (with I and V present vestigially), while dinosaurs are known from Triassic fossils to have a grasping, Raking hand with manual digits I-II-III, digits IV and V being greatly reduced in many Triassic forms. This is still a pivotal issue and an egregious omission for this book, with important papers by Čapek et al. [32Čapek DB, Metschler BT, Müller GB. Thumbs down: A molecular-morphogenetic approach to avian digit homology. J Exper Zool, B. Mol Devel Evol 2013; 322: 1-1223.] and others [19Feduccia A. Riddle of the Feathered Dragons 2012.], not listed in this book. For most paleontologists the accepted view is that there was a homeotic shift that occurred somewhere “between Allosaurus and birds” (whatever that means?) resulting in avian manual condensations II-III-IV transforming into digits I-II-III. What could have possibly caused this mysterious and unique transition [16Feduccia A. Fantasy vs Reality: A critique of Smith et al.’s Bird Origins. Open J Orn 2016; 9: 14-38.
[http://dx.doi.org/10.2174/1874453201609010014] ]?
One can now ask if the newly designated Pennaraptora (oviraptorosaurids, troodontids and dromaeosaurids), along with birds, represents a clade distinctive from true theropods, with manual digits II-III-IV [15James F, Pourtless JA. Cladistics and the origin of birds: A review and two new analyses. Orn Monographs 2009; 66: 1-78.
[http://dx.doi.org/10.1525/om.2009.66.1.1] , 16Feduccia A. Fantasy vs Reality: A critique of Smith et al.’s Bird Origins. Open J Orn 2016; 9: 14-38.
[http://dx.doi.org/10.2174/1874453201609010014] ]? If so, the current phylogeny advocated by Chiappe and Meng is topsy-turvy.
CONCLUSION
At a time when all major aspects of the dino–bird nexus are claimed to be known, scientists from the British Museum and Cambridge University published a paper in Nature [33Baron MG, Norman DB, Barrett PM. A new hypothesis of dinosaur relationships and early dinosaur evolution. Nature 2017; 543: 501-6.
[http://dx.doi.org/10.1038/nature21700] ] challenging the most basic tenets of deep dinosaur phylogeny. The authors conclude that theropods and ornithischians form a clade, the Ornithoscelida, and place the sauropodomorphs with the earlier and primitive herrerasaurs. Whether true of not, this paper should provide a strong dose of apprehension concerning phylogenetic certitude displayed in Birds of Stone. We are clearly a long way from understanding the complicated phylogeny of archosaurs, dinosaurs and birds.
The excitement generated by the Chinese fossil discoveries has parallels to the American “bone wars” of the latter 19th century, with paleontologists from all over the world eager to get their names on descriptive papers of newly discovered fossils. In the flurry of activity since the Chinese discoveries, the American Museum and its associates (notably Norell, Chiappe, and Brusatte) have made it to the top of the heap, largely usurping the popular exposition of these fossils, resulting in the perception that there is no appreciable deviation from their monolithic vision of the origins of birds, feathers and avian flight. At a time when morphological definitions of taxa have been banished and almost everything about bird origins is thought to be known, one could ask why the controversy continues? Current research appears now to be directed towards verifying what is already thought to be well known in the context of the cladogram, while contrary views are summarily rejected and not even acknowledged.
Birds of Stone adheres to tenets of the brave new world of postmodern paleornithology in methodology and certitude. Ascertaining phylogeny from vertebrate fossils has plagued paleontologists for decades, ranging from phenetics (overall similarity) that was supplanted by pure Hennigian methods (matching of carefully selected shared, derived characters), but now transformed into a new mechanical methodology, involving large numbers of characters (up to 1,389), selected by human hands and dealt with by computers, once again yielding a measure of overall phenetic similarity. An animal is whatever the computer-generated cladogram indicates, with no explanation of defining morphological characters. The most egregious result historically has been a clade of loons and grebes (known by whole genome analyses to be unrelated), and a clade of flightless ratites, having evolved flightlessness only once, deep in the Cretaceous and distributed to disparate continents by vicariance biogeography [34Cracraft J. Avian evolution, Gondwana biogeography and the Cretaceous-Tertiary mass extinction event. Proc Biol Sci 2001; 268: 459-69.
[http://dx.doi.org/10.1098/rspb.2000.1368] ]. Considered a triumph of the Hennigian era, it is “A textbook example of Gondwanan vicariance. . . dismantled [35Mayr G. Avian evolution: The fossil record of birds and its paleobiological significance 2017.]”. Other casualties may now include the conservation movement, now faced with proposals of up to 14,000 species of birds, up from the classic 9,000. Why worry about bird extinctions when there are so many new species by taxonomic inflation?
Chiappe and Meng do a credible job of portraying the current paleontological consensus, carefully following the basic axioms [36Brusatte SL. A Mesozoic aviary. Science 2017; 355: 792-4.
[http://dx.doi.org/10.1126/science.aal2397] ], including the evolution of feathers before flight, the ground-up origin of flight, the widespread occurrence of protofeathers, and biomechanical reconstructions based on the view that these forms are all descended from obligately bipedal theropods. However, most of these explanations require invoking massive exaptations to justify their special pleading. Lack of interest in key characters, abandonment of morphological definitions of taxa, and conflation of early birds and true theropods have combined to create a difficult arena for exchange of ideas between factions. Interestingly, if pennaraptorans (oviraptorosaurs, dromaeosaurs and troodontids) are nested within Aves, as many now believe, non-parsimonious explanations, such as ground-up flight, feathers before flight, and massive non-Darwinian exaptations are unnecessary. The innumerable unanswered questions and problems plaguing the current popular view of the origin of birds and avian flight demand that the issue remain an open inquiry. Are all the Mesozoic creatures with avian pennaceous feathers, including anchiornithids, oviraptorosaurs and microraptors really the hidden birds of China?
As Sir Karl Popper (1963) emphasized: “It is easy to obtain confirmations, or verifications, for nearly every theory; if we look for confirmations. [37Popper K. Conjectures and refutations: The growth of scientific knowledge 1963.]” One can ask with respect to bird origins, are we miles ahead of the data and simply trying to confirm what is already thought to be fact?
Popular consensus hypotheses usually change at a glacial pace, it is said, one funeral at a time. But, given the glaring problems with the current dogma on bird origins, hopefully new forward-thinking workers devoid of time-honored baggage will explore new and better explanations for the current data. In the meantime, the field will no doubt continue as it has: Verify! Verify!
CONSENT FOR PUBLICATION
Not applicable
CONFLICT OF INTEREST
The author declares no conflict of interest, financial or otherwise.
ACKNOWLEDGEMENTS
For helpful discussions and editorial comments I thank D. Burnham, F. James, T. Lingham-Soliar and J. Ruben. Lingham-Soliar and Burnham kindly provided the images for Figures 1 and 2.
REFERENCES
| [1] | Chiappe LM. Glorified Dinosaurs: The Origin and Early Evolution of Birds 2007. |
| [2] | Chiappe LM, Meng Q. Birds of Stone, Baltimore, MD, Johns Hopkins University Press 2016; pp 294 |
| [3] | Lingham-Soliar T. The evolution of the feather: Sinosauropteryx, a colourful tail. J Orn 2011; 152: 567-77. [http://dx.doi.org/10.1007/s10336-010-0620-y] |
| [4] | Lingham-Soliar T. The dinosaurian origin of feathers: Perspective from dolphin (Cetacea) collagen fibers. Naturwissenschaften 2003; 90: 563-7. [http://dx.doi.org/10.1007/s00114-003-0483-7] |
| [5] | Lingham-Soliar T. The evolution of the feather: Sinosauropteryx, life, death and preservation of an alleged feathered dinosaur. J Orn 2012; 153: 699-711. [http://dx.doi.org/10.1007/s10336-011-0787-x] |
| [6] | Lingham-Soliar T, Feduccia A, Wang X. A new Chinese specimen indicates that ‘protofeathers’ in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibers. Proc Royal Soc Long B. Biol Sci 2007; 274: 1823-9. [http://dx.doi.org/10.1098/rspb.2007.0352] |
| [7] | Lingham-Soliar T, Murugan N. A new helical crossed-fibre structure of β-Keratin in flight feathers and Its biomechanical implications. PLoS One 2013; 8(6): e65849. [http://dx.doi.org/10.1371/journal.pone.0065849] |
| [8] | Lingham-Soliar T. The evolution of the feather: Scales on the tail of Sinosauropteryx and an interpretation of the dinosaur’s opisthotonic posture. J Orn 2013; 154: 455-63. [http://dx.doi.org/10.1007/s10336-012-0910-7] |
| [9] | Lingham-Soliar T. Rare soft tissue preservation showing fibrous structures in an ichthyosaur from the Lower Lias (Jurassic) of England. Proc Biol Sci 1999; 266: 2367-73. [http://dx.doi.org/10.1098/rspb.1999.0933] |
| [10] | Vincent P, Allemand R, Taylor PD, Suan G, Maxwell EE. New insights on the systematics, palaeoecoloty and palaeobiology of a plesiosaurian with soft tissue preservation from the Toarcian of Holzmaden, Germany. Sci Nat 2017; 104: 51-00. [http://dx.doi.org/10.1007/s00114-017-1472-6] |
| [11] | Xu X, Zhou Z, Wang X, Kuang X, Zhang F, Du X. Four-winged dinosaurs from China. Nature 2003; 421: 335-40. [http://dx.doi.org/10.1038/nature01342] |
| [12] | Gong E-P, Martin LD, Burnham DA, Falk AF, Hou L-h. A new species of Microraptor from the jehol biota of northeastern china. Palaeoworld 2012; 21: 81-91. [http://dx.doi.org/10.1016/j.palwor.2012.05.003] |
| [13] | Feduccia A. Bird origins anew. Auk 2013; 130(1): 1-12. [http://dx.doi.org/10.1525/auk.2013.130.1.1] |
| [14] | Burnham DA, Feduccia A, Martin LD, Falk AR. Tree climbing: A fundamental avian adaptation. J Syst Palaeontology 2011; 9: 103-7. [http://dx.doi.org/10.1080/14772019.2010.522201] |
| [15] | James F, Pourtless JA. Cladistics and the origin of birds: A review and two new analyses. Orn Monographs 2009; 66: 1-78. [http://dx.doi.org/10.1525/om.2009.66.1.1] |
| [16] | Feduccia A. Fantasy vs Reality: A critique of Smith et al.’s Bird Origins. Open J Orn 2016; 9: 14-38. [http://dx.doi.org/10.2174/1874453201609010014] |
| [17] | Maryańska T, Osmólska H, Wolsan M. Avialan status for Oviraptorosauria. Acta Palaeontol Pol 2002; 47: 97-116. |
| [18] | Feduccia A, Czerkas SA. Testing the neoflightless hypothesis: Propatagium reveals flying ancestry of oviraptorosaurs. J Ornithol 2015; 156(4): 1067-74. [http://dx.doi.org/10.1007/s10336-015-1190-9] |
| [19] | Feduccia A. Riddle of the Feathered Dragons 2012. |
| [20] | Goldberg EE, Igi’c B. On phylogenetic tests of irreversible evolution. Evolution 2008; 62: 2727-41. [http://dx.doi.org/10.1111/j.1558-5646.2008.00505.x] |
| [21] | Chiappe LM. The dinosaur conspiracy. Bioscience 2012; 62(8): 770-2. [http://dx.doi.org/10.1525/bio.2012.62.8.11] |
| [22] | Falk AR, Kaye TG, Zhou Z, Burnham DA. Laser fluorescence illuminates the soft tissue and life habits of the Early Cretaceous bird Confuciusornis. PLoS One 2016; 11(12): e0167284. [http://dx.doi.org/10.1371/journal.pone.0167284] |
| [23] | Zheng X, O’Conner JK, Wang X, et al. Exceptional preservation of soft tissue in a new specimen of Eoconfuciusornis and its biological implications. Natl Sci Rev 2017; 4: 441-52. [doi: 10.1093/nst/nwx004]. [http://dx.doi.org/10.1093/nsr/nwx004] |
| [24] | Wang X, Pittman M, Zheng X, Kaye TG. Anchiornis soft tissues revealed by laser-stimulated fluorescence. Nat Commun 2017; 8: 14576a. [http://dx.doi.org/10.1038/ncomms14576] |
| [25] | Padian KL, Chiappe LM. The origin of birds and their flight. Sci Am 1998; 279: 38-47. [http://dx.doi.org/10.1038/scientificamerican0298-38] |
| [26] | Chiappe LM, Ji S-A, Qiang J, Norell MA. Anatomy and systematics of the Confuciusornithidae (Theropoda, Aves) from the late Mesozoic of northeastern China. 1999. |
| [27] | Olson SL. Review of anatomy and systematics of the confuciusornithidae (Theropoda: Aves) from the late mesozoic of China. Auk 2000; 117(3): 836-9. |
| [28] | Evangelista D, Cam S, Huynh T, et al. Shifts in stability and control effectiveness during evolution of Paraves support aerial maneuvering hypotheses for flight origins. PeerJ 2014; 2: e632. [http://dx.doi.org/10.7717/peerj.632] |
| [29] | Chatterjee S. The Rise of Birds 2nd ed. 2015. |
| [30] | Alexander DE. On the Wing: Insects, pterosaurs, birds, bats and the evolution of animal flight 2015. |
| [31] | Dudley R, Byrnes G, Yanoviak SP, Borrell B, Brown RM, McGuire JA. Gliding and the functional origins of flight: biomechanical novelty or necessity? Annu Rev Ecol Evol Syst 2007; 38: 179-201. [http://dx.doi.org/10.1146/annurev.ecolsys.37.091305.110014] |
| [32] | Čapek DB, Metschler BT, Müller GB. Thumbs down: A molecular-morphogenetic approach to avian digit homology. J Exper Zool, B. Mol Devel Evol 2013; 322: 1-1223. |
| [33] | Baron MG, Norman DB, Barrett PM. A new hypothesis of dinosaur relationships and early dinosaur evolution. Nature 2017; 543: 501-6. [http://dx.doi.org/10.1038/nature21700] |
| [34] | Cracraft J. Avian evolution, Gondwana biogeography and the Cretaceous-Tertiary mass extinction event. Proc Biol Sci 2001; 268: 459-69. [http://dx.doi.org/10.1098/rspb.2000.1368] |
| [35] | Mayr G. Avian evolution: The fossil record of birds and its paleobiological significance 2017. |
| [36] | Brusatte SL. A Mesozoic aviary. Science 2017; 355: 792-4. [http://dx.doi.org/10.1126/science.aal2397] |
| [37] | Popper K. Conjectures and refutations: The growth of scientific knowledge 1963. |