- Home
- About Journals
-
Information for Authors/ReviewersEditorial Policies
Publication Fee
Publication Cycle - Process Flowchart
Online Manuscript Submission and Tracking System
Publishing Ethics and Rectitude
Authorship
Author Benefits
Reviewer Guidelines
Guest Editor Guidelines
Peer Review Workflow
Quick Track Option
Copyediting Services
Bentham Open Membership
Bentham Open Advisory Board
Archiving Policies
Fabricating and Stating False Information
Post Publication Discussions and Corrections
Editorial Management
Advertise With Us
Funding Agencies
Rate List
Kudos
General FAQs
Special Fee Waivers and Discounts
- Contact
- Help
- About Us
- Search




The Open Ornithology Journal
(Discontinued)
ISSN: 1874-4532 ― Volume 13, 2020
Influence of Disturbance on Avian Communities in Agricultural Conservation Buffers in Mississippi, USA
Heidi L. Adams1, *, L. Wes Burger, Jr.2, Sam Riffell2
Abstract
Introduction:
Periodic disturbance of agricultural conservation buffers is required to maintain early successional plant communities for grassland birds. However, a disturbance may temporarily reduce the availability of vegetation cover, food, and nesting sites in a buffer.
Objective:
Our objective was to determine how the type of disturbance (i.e., prescribed burning, light disking) and time since the last disturbance event in agricultural conservation buffers influence the grassland bird community.
Methods:
Data collected during line-transect surveys conducted in 46 agricultural conservation buffers in northeast Mississippi during the 2007-2009 breeding seasons (May-early August) demonstrate periodic disturbance through prescribed burning and light disking does not influence breeding bird diversity or density in the buffers.
Results:
Density of Dickcissels (Spiza americana), Red-winged Blackbirds (Agelaius phoeniceus), and Indigo Buntings (Passerina cyanea) did not differ in the buffers regardless of the type of or time since disturbance.
Conclusion:
Large effect sizes, however, indicate a potential type two error resulting from this conclusion. Thus, based on relative effect sizes, avian density in undisturbed buffers may be greater than in buffers during their first growing season post-disturbance. Relative effect sizes among estimates also indicate disturbance, namely prescribed burning, may lead to greater densities of breeding birds in agricultural conservation buffers. Though disturbance may initially reduce avian density, it is necessary to maintain long-term early-successional herbaceous habitat in agricultural conservation buffers.
Article Information

Identifiers and Pagination:
Year: 2019Volume: 12
First Page: 16
Last Page: 29
Publisher Id: TOOENIJ-12-16
DOI: 10.2174/187445320191200016
Article History:
Received Date: 07/07/2019Revision Received Date: 19/10/2019
Acceptance Date: 19/11/2019
Electronic publication date: 31/12/2019
Collection year: 2019
open-access license: This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International Public License (CC-BY 4.0), a copy of which is available at: (https://creativecommons.org/licenses/by/4.0/legalcode). This license permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
* Address correspondence to this author at the School of Agricultural Sciences and Forestry, Louisiana Tech University, P. O. Box 10138, Ruston, LA 71270, USA; Tel: 318-257-2947; E-mail: hadams@latech.edu
| Open Peer Review Details | |||
|---|---|---|---|
| Manuscript submitted on 07-07-2019 |
Original Manuscript | Influence of Disturbance on Avian Communities in Agricultural Conservation Buffers in Mississippi, USA | |
1. INTRODUCTION
Global agricultural crop production has changed greatly in the last 100 years. With technological advances, such as mechanization, inorganic fertilizers, and chemical use, there has been a disassociation between agriculture and natural resources [1Smeding FW, Joenje W. Farm-nature plan: Landscape ecology based on farm planning. Landsc Urban Plan 1999; 46: 109-15.
[http://dx.doi.org/10.1016/S0169-2046(99)00052-3] -3Liira J, Aavik T, Parrest O, Zobel M. Agricultural sector, rural environment and biodiversity in the central and eastern European EU member states. AGD Landsc Environ 2008; 2: 46-64.]. After World War II, for instance, European cereal production increased in an effort to be more self-sufficient [4Robinson RA, Sutherland WJ. Post-war changes in arable farming and biodiversity in Great Britain. J Appl Ecol 2002; 39: 157-76.
[http://dx.doi.org/10.1046/j.1365-2664.2002.00695.x] ]. This increase led to the overproduction of crops and a reduction in landscape biodiversity [4Robinson RA, Sutherland WJ. Post-war changes in arable farming and biodiversity in Great Britain. J Appl Ecol 2002; 39: 157-76.
[http://dx.doi.org/10.1046/j.1365-2664.2002.00695.x] ]. In the United States, agricultural cropland covers nearly 318 million ha, constituting 14% of the country’s total land use [5United States Department of Agriculture. National Agricultural Statistics Service [accessed 2018 May 22] Available from: https://www.nass.usda.gov/]. These agricultural systems are often characterized by large, monocultural fields that have fragmented or replaced natural ecosystems important to native flora and fauna [6Benton TG, Vickery JA, Wilson JD. Farmland biodiversity: Is habitat heterogeneity the key? Trends Ecol Evol 2003; 18: 182-8.
[http://dx.doi.org/10.1016/S0169-5347(03)00011-9] , 7Burger LW Jr, Smith MD, Barbour PJ, Hamrick R. Grassland bird response to agricultural field borders. Forest Wildl Res Center Res Adv 2005; 8: 1-4.].
Habitat loss associated with agricultural conversion and intensification is the greatest threat to declining bird populations, particularly grassland birds [8Green RE, Cornell SJ, Scharlemann JPW, Balmford A. Farming and the fate of wild nature. Science 2005; 307(5709): 550-5.
[http://dx.doi.org/10.1126/science.1106049] [PMID: 15618485] ]. In some regions of North America, there has been a loss of up to 99.9% of natural grassland ecosystems since the birth of industrialized agriculture [9Samson F, Knopf K. Prairie conservation in North America. Bioscience 1994; 44: 418-21.
[http://dx.doi.org/10.2307/1312365] ]. In turn, grassland birds have shown steeper and more consistent declines than any other breeding bird guild [10Knopf FL. Avian assemblages on altered grasslands. Stud Avian Biol 1994; 15: 247-57., 11Donal PF, Gree RE, Heath MF. Agricultural intensification and the collapse of Europe’s farmland bird populations. Proc Biol Sci 2001; 268(1462): 25-9.
[http://dx.doi.org/10.1098/rspb.2000.1325] [PMID: 12123294] ]. Examples of species that experienced population declines in the United States in recent decades include Dickcissel (Spiza americana), Eastern (Sturnella magna) and Western Meadowlark (Sturnella neglecta), Bobolink (Dolichonyx oryzivorus), and Northern Bobwhite (Colinus virginianus) [11Donal PF, Gree RE, Heath MF. Agricultural intensification and the collapse of Europe’s farmland bird populations. Proc Biol Sci 2001; 268(1462): 25-9.
[http://dx.doi.org/10.1098/rspb.2000.1325] [PMID: 12123294] ]. Similar trends have also been found in Europe. From 1970 to 1990, for instance, there was a negative relationship between European avian population sizes and agricultural intensification, especially in Western Europe [12Pedroli GBM, Meiner A. Landscapes in transition: An account of 25 years of land cover change in Europe. Publications Office of the European Union, EEA Report No 10/2017 2017.]. As of 2012, 25% of land cover in 39 European countries, totaling 1,471,684 km2, was classified as arable land and permanent crops [12Pedroli GBM, Meiner A. Landscapes in transition: An account of 25 years of land cover change in Europe. Publications Office of the European Union, EEA Report No 10/2017 2017.].
A reversion to less intensive agriculture is often not feasible, thus conservation practices that incorporate natural areas for birds and other wildlife into existing agricultural systems may be the best alternative. This is especially true in field margins adjacent to more competitive vegetation, such as trees and woody shrubs [13Barbour PJ, Martin SW, Burger W. Estimating economic impact of conservation field borders on farm revenue. Crop Manag 2007; 6
[http://dx.doi.org/10.1094/CM-2007-0614-01-RS] ]. Agricultural conservation buffers may be integrated easily into production systems and produce many conservation benefits with minimal changes in primary land use. When buffers were established around crop fields in Sweden, for instance, there was a 30% increase in Eurasian Skylark (Alauda avensis) density relative to fields without buffers [14Josefsson J, Berg A, Hiron M, Pärt T, Eggers S. Grass buffer strips benefit invertebrate and breeding skylark numbers in a heterogeneous agricultural landscape. Agric Ecosyst Environ 2013; 181: 101-7.
[http://dx.doi.org/10.1016/j.agee.2013.09.018] ]. In the United States, breeding grassland bird densities in buffered crop fields were greater compared to those crop fields without buffers [15Evans KO, Burger LW Jr, Riffell S, Smith MD. Assessing multiregion avian benefits from strategically targeted agricultural buffers. Conserv Biol 2014; 28(4): 892-901.
[http://dx.doi.org/10.1111/cobi.12311] [PMID: 24779636] ]. Species-specific density differences included Northern Bobwhite (85-109% greater), Dickcissel (85-120% greater), and Field Sparrow (Spizella pusilla; 58-106% greater) [15Evans KO, Burger LW Jr, Riffell S, Smith MD. Assessing multiregion avian benefits from strategically targeted agricultural buffers. Conserv Biol 2014; 28(4): 892-901.
[http://dx.doi.org/10.1111/cobi.12311] [PMID: 24779636] ]. Buffers can provide many other benefits, as well. Examples include serving as corridors to facilitate wildlife movement across the landscape, promoting populations of beneficial insects (e.g., pollinators), reducing soil erosion and agrochemical runoff, and increasing farm-level biodiversity [2Marshall EJP, Moonen AC. Field margins in northern Europe: Their functions and interactions with agriculture. Agric Ecosyst Environ 2002; 89: 5-21.
[http://dx.doi.org/10.1016/S0167-8809(01)00315-2] , 16Lovell ST, Sullivan WC. Environmental benefits of conservation buffers in the United States: evidence, promise, and open questions. Agric Ecosyst Environ 2006; 112: 249-60.
[http://dx.doi.org/10.1016/j.agee.2005.08.002] , 17Dollar JG, Riffell SK, Burger LW Jr. Effects of managing semi-natural grassland buffers on butterflies. J Insect Conserv 2013; 7: 577-90.
[http://dx.doi.org/10.1007/s10841-012-9543-7] ].
While buffers may provide many ecological benefits, they are often established with a specific, predetermined management goal [18Olson DM, Wäckers FL. Management of field margins to maximize multiple ecological services. J Appl Ecol 2007; 44: 13-21.
[http://dx.doi.org/10.1111/j.1365-2664.2006.01241.x] ]. Buffers established with the specific objective of promoting grassland bird populations must be periodically disturbed to maintain them as early-successional habitat [7Burger LW Jr, Smith MD, Barbour PJ, Hamrick R. Grassland bird response to agricultural field borders. Forest Wildl Res Center Res Adv 2005; 8: 1-4., 19McCoy TD, Kurzejeski EW, Burger LW Jr, Ryan MR. Effects of conservation practice, mowing, and temporal changes on vegetation structure on Conservation Reserve Program fields in northern Missouri. Wildl Soc Bull 2001; 29: 979-87.]. Disturbance, such as prescribed burning and light disking, will not only prevent woody plant encroachment, but also enhance herbaceous structural diversity, reduce vegetation density, decrease litter cover, increase the abundance of bare ground, and promote plant diversity [20Rodenhouse NL, Best LB, O’Connor RJ, Bollinger EK. Effects of agricultural practices on farmland structures. In: Martin TE, Finch DM, Eds. Ecology and management of Neotropical migratory birds: A synthesis and review of critical issues 1995; 269-93.-23Fuhlendorf SD, Harrell WC, Engle DM, Hamilton RG, Davis CA, Leslie DM Jr. Should heterogeneity be the basis for conservation? Grassland bird response to fire and grazing. Ecol Appl 2006; 16(5): 1706-16.
[http://dx.doi.org/10.1890/1051-0761(2006)016[1706:SHBTBF]2.0.CO;2] [PMID: 17069365] ]. In turn, the ability of buffers to support breeding grassland birds is enhanced. When patches of Scottish buffers were cut, for instance, Yellowhammer (Emberiza citrinella) summer foraging activity was greater than in uncut patches [24Douglas DJT, Vickery JA, Benton TG. Improving the value of field margins as foraging habitat for farmland birds. J Appl Ecol 2009; 46: 353-62.
[http://dx.doi.org/10.1111/j.1365-2664.2009.01613.x] ]. In North Dakota, prescribed fire in mixed-grass prairies prevented woody plant encroachment without negatively influencing nest survival of the Clay-colored (Spizella pallida) and Savannah sparrow (Passerculus sandwichensis) [25Murphy RK, Shaffer TL, Grant TA, Derrig JL, Rubin CS, Kerns CK. Sparrow nest survival in relation to prescribed fire and woody plant invasion in a northern mixed-grass prairie. Wildl Soc Bull 2017; 41: 442-52.
[http://dx.doi.org/10.1002/wsb.780] ]. The use of fire as a management tool, however, has declined in recent decades. In areas heavily influenced by humans, issues regarding smoke and fire management are of great concern. Agricultural systems fragmented by roads, developed areas, crop fields, and pastures may require fire-free buffers and smoke management [26Engstrom RT, Vickery PD, Perkins DW, Shriver WG. Effects of fire regime on birds in southeastern pine savannas and native prairies. Stud Avian Biol 2005; 30: 147-60.]. Additionally, there may be public concerns about potential negative effects fire may have on air quality in and around a burn area [27Haikerwal A, Reisen F, Sim MR, et al. Impact of smoke from prescribed burning: Is it a public health concern? J Air Waste Manag Assoc 2015; 65(5): 592-8.
[http://dx.doi.org/10.1080/10962247.2015.1032445] [PMID: 25947317] ].
Time since disturbance may also influence grassland bird diversity and density in buffers. For instance, immediately following disturbance, a buffer may have short and sparse grass cover, few forbs, and minimal litter cover, thus providing little in regards to nesting and foraging habitat for breeding grassland birds [22Reinking DL. Fire regimes and avian responses in the central tallgrass prairie. Stud Avian Biol 2005; 30: 116-26.]. In an east-central North Dakota mixed-grass prairie, densities of Bobolink, Western Meadowlark, and Grasshopper Sparrow (Ammodramus savannarum) decreased immediately following a burn [28Johnson DH. Effects of fire on bird populations in mixed-grass prairie. In: Knopf FL, Samson FB, Eds. Ecology and conservation of Great Plains vertebrates 1997; 181-206.
[http://dx.doi.org/10.1007/978-1-4757-2703-6_8] ]. Between 2 and 8 years later, however, this same area had an increasing abundance of grasses, forbs, shrubs, and litter cover [28Johnson DH. Effects of fire on bird populations in mixed-grass prairie. In: Knopf FL, Samson FB, Eds. Ecology and conservation of Great Plains vertebrates 1997; 181-206.
[http://dx.doi.org/10.1007/978-1-4757-2703-6_8] ]. Thus, fire influences short and long-term habitat suitability for birds. Specific response to time after burn will vary among species and regions in relation to precipitation regimes and soil fertility.
For these reasons, the use of alternative types of disturbance, such as light disking, must be explored to determine which management methods are most effective for supporting grassland bird populations in buffers. Thus, our objective was to determine the effect of periodic disturbance (e.g., prescribed burning and light disking) on the breeding grassland bird diversity and density in agricultural conservation buffers during the avian breeding seasons of 2007-2009. We hypothesized prescribed burning would result in greater diversity and density of breeding grassland birds relative to light disking. The results of our study will aid private land owners and land managers in selecting the most appropriate disturbance tool for areas selected to be early-successional buffer habitat in agricultural systems to support grassland bird populations.
2. METHODS
2.1. Data Collection
We collected data at a privately-owned farm in northeast Mississippi, USA, which consisted of 2,104 ha, 486 ha of which were used for row crop production and 587 ha that were used for a cattle operation (Fig. 1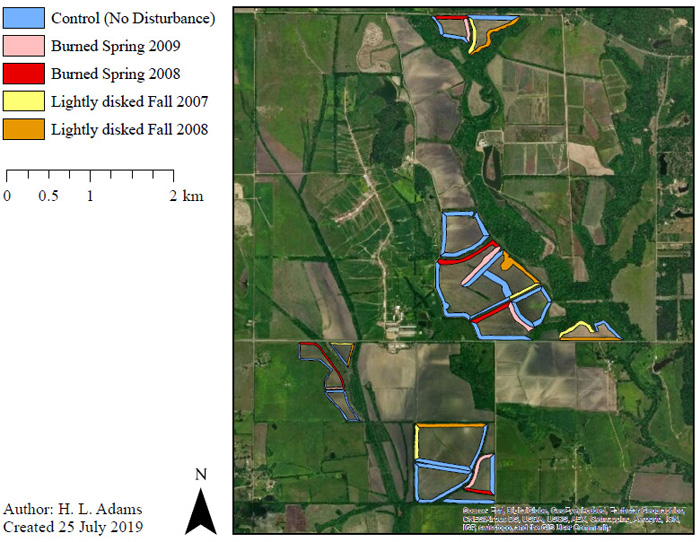 ). In spring 2005, 79 ha of this property were enrolled in the Conservation Reserve Program’s Conservation Practice 33 (CP33: Habitat Buffers for Upland Birds). Based on the enrollment criteria for this practice, agricultural conservation buffers 9.1-36.5 m wide are established around the perimeter of crop production fields with native warm-season grasses and forbs. Producers implementing CP33 on their land receive a cost-share to offset establishment costs, as well as monetary incentives and annual rental payments to compensate for lost opportunity costs [29Burger LW Jr, McKenzie D, Thackston R, Demaso SJ. The role of farm policy in achieving large-scale conservation: Bobwhite and buffers. Wildl Soc Bull 2006; 34: 986-93.
). In spring 2005, 79 ha of this property were enrolled in the Conservation Reserve Program’s Conservation Practice 33 (CP33: Habitat Buffers for Upland Birds). Based on the enrollment criteria for this practice, agricultural conservation buffers 9.1-36.5 m wide are established around the perimeter of crop production fields with native warm-season grasses and forbs. Producers implementing CP33 on their land receive a cost-share to offset establishment costs, as well as monetary incentives and annual rental payments to compensate for lost opportunity costs [29Burger LW Jr, McKenzie D, Thackston R, Demaso SJ. The role of farm policy in achieving large-scale conservation: Bobwhite and buffers. Wildl Soc Bull 2006; 34: 986-93.
[http://dx.doi.org/10.2193/0091-7648(2006)34[986:TROFPI]2.0.CO;2] ]. Buffers enrolled in CP33 are required to be disturbed to maintain them as early-successional habitat specifically for Northern Bobwhite and potentially other grassland birds [29Burger LW Jr, McKenzie D, Thackston R, Demaso SJ. The role of farm policy in achieving large-scale conservation: Bobwhite and buffers. Wildl Soc Bull 2006; 34: 986-93.
[http://dx.doi.org/10.2193/0091-7648(2006)34[986:TROFPI]2.0.CO;2] ].

After CP33 enrollment, buffers measuring 18.2 or 36.5 m wide were then established, surrounding the perimeter of 14 crop production fields at the farm. These buffers included a mix of native warm-season grasses [e.g., big bluestem (Andropogon gerardi), little bluestem (Schizachyrium scoparium), indiangrass (Sorghastrum nutans)] and forbs [e.g., partridge pea (Chamaecrista fasciculata), black-eyed susan (Rudbeckia hirta), maximilian sunflower (Helianthus maximiliani)].
We investigated 3 different disturbance treatments in agricultural conservation buffers at the study site: (1) Light disking in the fall (September-October), (2) Prescribed burning in the spring (March-April), and (3) No disturbance (control). Because data collection occurred during the avian breeding season (May-early August), confounding between disturbance type and breeding season did not influence the data. We randomly assigned 1 of the 3 disturbance treatments to each group of buffers surrounding the same field, with only 1 buffer/field group disturbed each year (14 buffer-bordered fields, 51 total buffers). Planted crops in the fields included a corn-soybean rotation or Bermuda grass (Cynodon dactylon) established for cattle forage. Data we collected during 2007 provided pre-disturbance information. We documented vegetation and bird responses to disturbance during the first (2008 and 2009) and second (2009) growing seasons post-disturbance. Because this study concluded prior to 2010, we considered buffers disturbed after the 2009 breeding season in-field controls.
We used distance sampling techniques to estimate breeding bird density as it incorporates a decreasing detection probability with increasing distance from an observer [30Buckland ST, Anderson DR, Burnham KP, Laake JL, Borchers DL, Thomas L. Introduction to distance sampling 2001.]. We established 200-m long line transects that ran parallel to a buffer’s long axis in 46 of the agricultural conservation buffers at the study site [5 buffers were too short to accommodate a 200-m line transect; (Fig. 2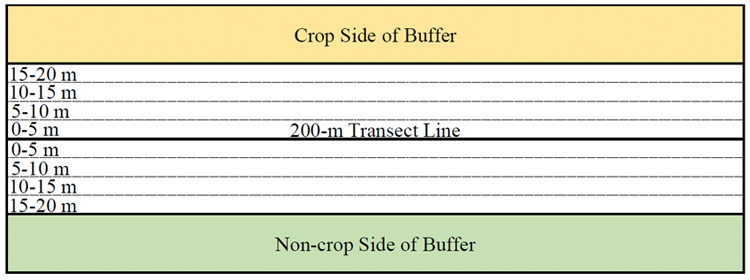 )]. Strip transect surveys were conducted by a single observer (HLA) 0530-1000 (central standard time) on mornings with no precipitation and wind speeds less than 24 km/hour [31Smith MD, Barbour PJ, Burger LW Jr, Dinsmore SJ. Breeding bird abundance and diversity in agricultural field borders in the Black Belt Prairie of Mississippi. Proc Annu Conf Southeast Assoc Fish Wildl Agencies 2005; 43-56.]. Fixed-width transect surveys were conducted at a travel rate of 10 m/min, during which the observer recorded bird detections in 1 of 4 distance bands that together covered the width of the buffers: 0-5 m, 5-10 m, 10-15 m, and 15-20 m. Temperature, percent cloud coverage, and wind speed were recorded at the start of each transect survey. The observer visited each transect 6 times during each breeding season (twice monthly, May-early August), 2007-2009.
)]. Strip transect surveys were conducted by a single observer (HLA) 0530-1000 (central standard time) on mornings with no precipitation and wind speeds less than 24 km/hour [31Smith MD, Barbour PJ, Burger LW Jr, Dinsmore SJ. Breeding bird abundance and diversity in agricultural field borders in the Black Belt Prairie of Mississippi. Proc Annu Conf Southeast Assoc Fish Wildl Agencies 2005; 43-56.]. Fixed-width transect surveys were conducted at a travel rate of 10 m/min, during which the observer recorded bird detections in 1 of 4 distance bands that together covered the width of the buffers: 0-5 m, 5-10 m, 10-15 m, and 15-20 m. Temperature, percent cloud coverage, and wind speed were recorded at the start of each transect survey. The observer visited each transect 6 times during each breeding season (twice monthly, May-early August), 2007-2009.
 |
Fig. (2) Diagram of a 200-m transect used to estimate diversity and density of breeding birds in agricultural conservation buffers at a privately-owned farm in northeast Mississippi, USA, 2007-2009. |
2.2. Data Analyses
We estimated avian diversity in the agricultural conservation buffers using Shannon’s Diversity Index [32Shannon CE, Weaver W. The mathematical theory of communication 1963.], which is calculated as:
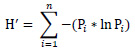 |
(1) |
where H’ is the Shannon’s Diversity Index value, n is the number of detected species, and Pi is the fraction of the observed sample composed of the ith species. We calculated diversity indices for the type of and time since disturbance using mean detections/ha for all birds detected during line-transect surveys.
To determine if disturbance influenced avian diversity at the buffer level, we compared mean avian diversity in buffers based upon variables reflecting year (2007, 2008, 2009) by treatment (burn, disk, control) using general linear models in SAS PROC MIXED [33SAS 9.4 product documentation. 2013.]. Because individual buffers of the same field were physically connected and adjacent to the same crop field, we included field as a random variable. To determine if disturbance influenced avian density at the field level, we compared the mean avian diversity of fields based on year by treatment variables using general linear models in SAS PROC MIXED, including year as a repeated measure [33SAS 9.4 product documentation. 2013.]. At both the buffer and field level, we further evaluated significant results using a Welch t-test. For all these statistical analyses, we used a significance level of α = 0.05.
We used conventional distance sampling techniques in Program Distance to estimate the detection function of all birds of any species in the agricultural conservation buffers based on the type of and time since disturbance [34Thomas L, Buckland ST, Rexstad EA, et al. Distance software: design and analysis of distance sampling surveys for estimating population size. J Appl Ecol 2010; 47(1): 5-14.
[http://dx.doi.org/10.1111/j.1365-2664.2009.01737.x] [PMID: 20383262] ]. Additionally, we estimated detection functions for species with more than 100 detections in the buffers (i.e., Dickcissel, Red-winged Black- bird, Indigo Bunting). Species with less than 100 detections provided insufficient data to generate a robust estimate of a detection function [30Buckland ST, Anderson DR, Burnham KP, Laake JL, Borchers DL, Thomas L. Introduction to distance sampling 2001.].
It is recommended that approximately 5-10% of distance data be right-truncated to increase model precision [30Buckland ST, Anderson DR, Burnham KP, Laake JL, Borchers DL, Thomas L. Introduction to distance sampling 2001.]. Because we recorded detections in 5-m distance bands rather than exact distances, truncation required censoring all or none of the detections in a given band. The 15-20 m band terminated at the buffer-crop ecotone at a transect’s crop side (i.e., field interior) and the buffer-field margin ecotone at a transect’s non-crop side (i.e., field exterior). Vegetation discontinuities associated with edges attracted birds, resulting in more detections in the outer distance bands (15-20 m) and a non-monotonically declining detection function with distance (Fig. 3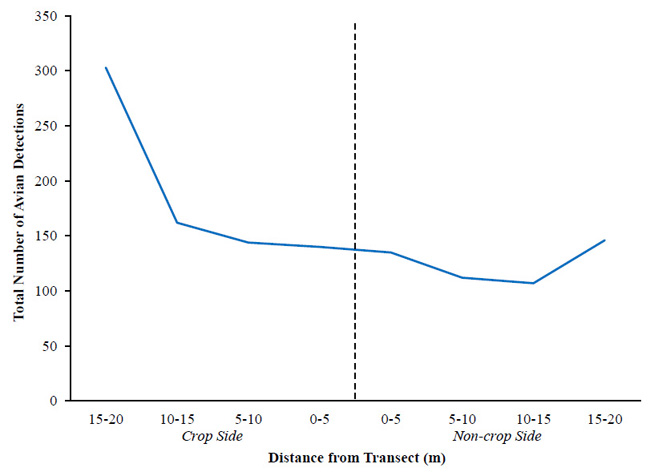 ). This violated an assumption of distance sampling, which states the probability of detecting an object; in this case, a bird, decreases with increasing distance from an observer [30Buckland ST, Anderson DR, Burnham KP, Laake JL, Borchers DL, Thomas L. Introduction to distance sampling 2001.]. Thus, we truncated detections in this band and estimated detection functions using detections in the first 3 distance bands (0-15 m). We used Akaike’s Information Criterion (AIC) to determine the best fit detection function from appropriate key functions (half-normal or uniform) with possible cosine or simple polynomial adjustment terms. Model selection was based on the least AIC value, goodness-of-fit, and detection probability.
). This violated an assumption of distance sampling, which states the probability of detecting an object; in this case, a bird, decreases with increasing distance from an observer [30Buckland ST, Anderson DR, Burnham KP, Laake JL, Borchers DL, Thomas L. Introduction to distance sampling 2001.]. Thus, we truncated detections in this band and estimated detection functions using detections in the first 3 distance bands (0-15 m). We used Akaike’s Information Criterion (AIC) to determine the best fit detection function from appropriate key functions (half-normal or uniform) with possible cosine or simple polynomial adjustment terms. Model selection was based on the least AIC value, goodness-of-fit, and detection probability.
Given that model selection results most frequently supported uniform detection probabilities at or near 1.0 and a relatively narrow detection width, we estimated bufferand year-specific avian density in the agricultural conservation buffers with respect to type of and time since disturbance by calculating mean number of avian detections/ha for each buffer during 2008 and 2009. We made these estimations using all avian detection data and not just the truncated data used in Program Distance analyses. We evaluated potential differences in avian density between control and disturbed buffers at the buffer and field level using general linear models in SAS PROC MIXED [33SAS 9.4 product documentation. 2013.]. Procedures for these analyses were similar to diversity analyses, again with α = 0.05.
3. RESULTS
We detected 26 avian species in the agricultural conservation buffers at the study site during the 2007-2009 breeding seasons (Table 1). Of these 26 species, 5 were grassland species, 6 were facultative grassland, 9 were edge species that use the field-forest ecotone, 3 were woodland species, and 3 were more commonly associated with urban or developed areas. Dickcissels (grassland, 427 detections), Red-winged Blackbirds (facultative grassland, 336 detections), and Indigo Buntings (edge, 252 detections) were the most frequently detected species in the agricultural conservation buffers.
 |
Fig. (3) The total number of birds detected during strip transect surveys in agricultural conservation buffers at a privately-owned farm in northeast Mississippi, USA, 2007-2009. |

3.1. Avian Diversity
There were no pre-treatment differences in mean avian diversity in 2007 among the agricultural conservation buffers (F4,34.9 = 0.65, P = 0.630). There was still no difference in diversity at the buffer level regardless of type of or time since disturbance in 2008 (F2,36 = 1.34, P = 0.274) and 2009 (F4,37.4 = 0.32, P = 0.864). In 2008, although relative effect sizes were large, -29% for burned buffers and -92% for disked buffers compared to control buffers suggesting a short-term reduction in avian diversity the first growing season post-disturbance, confidence intervals included zero (Fig. 4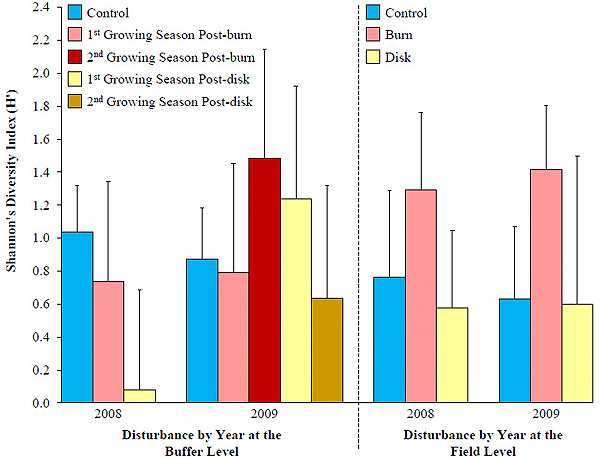 ). In 2009, effect sizes on diversity were 70% in burned buffers in their second growing season and 42% for disked buffers in their first growing season relative to controls, suggesting a positive response to disturbance, although confidence intervals again included 0.
). In 2009, effect sizes on diversity were 70% in burned buffers in their second growing season and 42% for disked buffers in their first growing season relative to controls, suggesting a positive response to disturbance, although confidence intervals again included 0.
At the field level in 2007, there were no pre-treatment differences in avian diversity (F2,11 = 0.69, P = 0.523). Implementation of disturbance treatments did not affect avian diversity in 2008 (F2,11 = 0.62, P = 0.557) and 2009 (F2,11 = 1.37, P = 0.294). Estimates of effect sizes (relative to control fields) were relatively large and in opposite directions for prescribed burning (2008, 69% increase in diversity; 2009, 124%) and light strip-disking (2008, 25% decrease in diversity; 2009, 5%).
3.2. Avian Density
All model selection results generated in Program Distance supported the uniform key function with a cosine adjustment to estimate detection functions regardless of the type of or time since disturbance (Table 2). Detection probabilities were also all 1.00 ± 0.00 for Dickcissels (Table 3) and Indigo Buntings (Table 4); Red-winged Blackbird detection probabilities were all 1.00 ± 0.00 except for buffers during the second growing season post-disking (0.50 ± 0.12; Table 5).


Results of Akaike's information criterion (AIC), detection function [f(0)], probability of detection (p), and percent coefficient of variation (%CV) for all birds detected in control and disturbed agricultural conservation buffers at a privately-owned farm in northeast Mississippi, USA, 2007-2009.

Results of Akaike's information criterion (AIC), detection function [f(0)], probability of detection (p), and percent coefficient of variation (%CV) for Dickcissels (Spiza americana) detected in control and disturbed agricultural conservation buffers at a privately-owned farm in northeast Mississippi, USA, 2007-2009.

Results of Akaike's information criterion (AIC), detection function [f(0)], probability of detection (p), and percent coefficient of variation (%CV) for Indigo Buntings (Passerina cyanea) detected in control and disturbed agricultural conservation buffers at a privately-owned farm in northeast Mississippi, USA, 2007-2009.

Results of Akaike's information criterion (AIC), detection function [f(0)], probability of detection (p), and percent coefficient of variation (%CV) for Red-winged Blackbirds (Agelaius phoeniceus) detected in control and disturbed agricultural conservation buffers at a privately-owned farm in northeast Mississippi, USA, 2007–2009.
At the buffer level in 2007, there was no difference in avian density among buffers prior to disturbance (F4,35.1 = 1.76, P = 0.160). Avian density in control and disturbed buffers did not differ in 2008 (F2,35.4 = 0.99, P = 0.383) and 2009 (F4,37.4 = 0.58, P = 0.680). In 2008, relative effect sizes on avian density were -26% for burned buffers in the first growing season and -68% for disked buffers in the first growing season (Fig. 5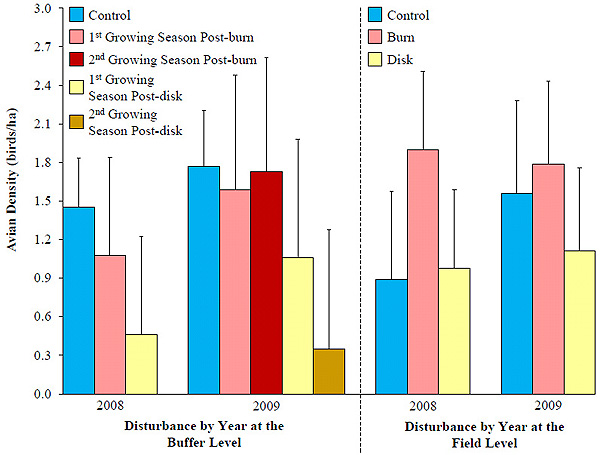 ), reflecting a short-term decline in density. However, confidence intervals included 0. In 2009, relative effect sizes on total avian density were -10% for buffer the first growing season post-burn and -2% the second season post-burn. Again, confidence intervals on effect sizes included 0.
), reflecting a short-term decline in density. However, confidence intervals included 0. In 2009, relative effect sizes on total avian density were -10% for buffer the first growing season post-burn and -2% the second season post-burn. Again, confidence intervals on effect sizes included 0.
At the field level, prior to disturbance, there was no difference in avian density among fields bordered by agricultural conservation buffers (F2,11 = 0.59, P = 0.572). After disturbance regimes had been implemented, total avian density did not differ among treatments in 2008 (F2,11 = 0.80, P = 0.475) and 2009 (F2,11 = 0.28, P = 0.752). Relative effect sizes for avian density were positive for prescribed burning (2008, 112%; 2009, 15%), whereas effect sizes on avian density were mixed for disked fields (2008, 10%; 2009, -29%). However, field level confidence intervals on effect sizes included 0 for years and treatments.

3.3. Dickcissels
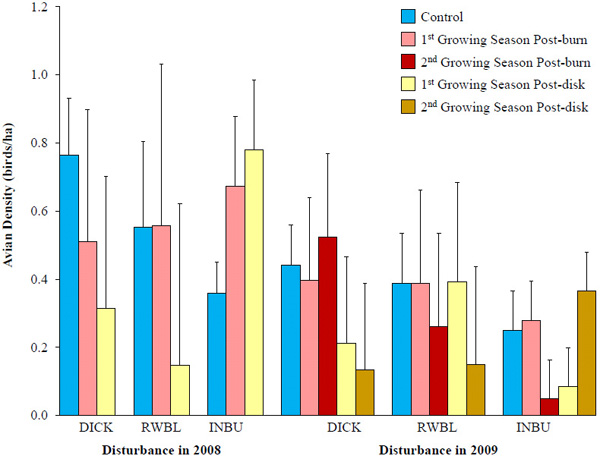
In 2007, there was no difference in Dickcissel density prior to disturbance (F4,38 = 0.98, P = 0.428). Furthermore, neither type of nor time since disturbance influenced Dickcissel density in buffers during 2008 (F2,37.7 = 0.81, P = 0.454) and 2009 (F4,38.9 = 0.50, P = 0.735). In 2008, Dickcissel density in buffers the first growing season post-burn was -33% relative to control buffers (Fig. 6 ). In 2009, relative effect size in Dickcissel density between these same buffers their second growing season post-burn and controls was 19%. Confidence intervals on effect sizes, however, included 0 for years and treatments.
). In 2009, relative effect size in Dickcissel density between these same buffers their second growing season post-burn and controls was 19%. Confidence intervals on effect sizes, however, included 0 for years and treatments.
At the field level in 2007, there was no difference in Dickcissel density (F2,11 = 0.01, P = 0.989). Post-disturbance Dickcissel density still did not differ among fields bordered by agricultural conservation buffers in 2008 (F2,11 = 0.09, P = 0.911) and 2009 (F2,11 = 0.54, P = 0.599). Effect sizes on mean Dickcissel density in fields with disking as the assigned treatment were -6% in 2008 and -3% in 2009 relative to control fields (Fig. 7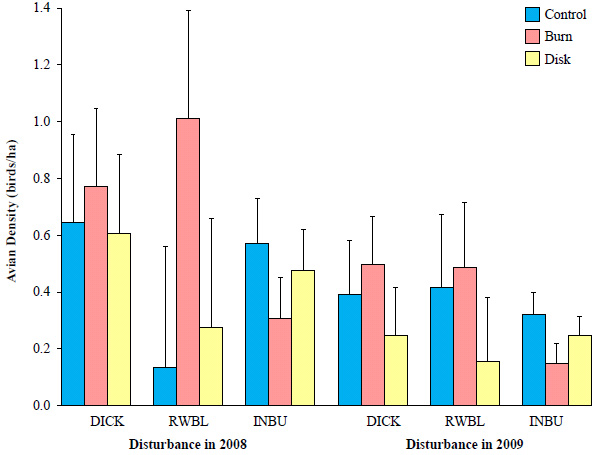 ). Conversely, the relative effect size in Dickcissel density between burned and control fields was 19% in 2008 and 27% in 2009. Though these results suggest a positive response to prescribed burning, confidence intervals included 0.
). Conversely, the relative effect size in Dickcissel density between burned and control fields was 19% in 2008 and 27% in 2009. Though these results suggest a positive response to prescribed burning, confidence intervals included 0.

3.4. Red-winged Blackbirds
In 2007, there was no difference in Red-winged Blackbird density in agricultural conservation buffers prior to disturbance (F4,34.7 = 1.43, P = 0.246). Blackbird density did not differ between disturbed and control buffers in 2008 (F2,34.8 = 0.40, P = 0.675) and 2009 (F4,36.5 = 0.21, P = 0.931). In 2008, there was a 73% difference in relative effect size between blackbird densities in buffers the first growing season post-disk and control buffers. Based on relative effect sizes in 2009, blackbird density in disked buffers compared to control buffers was -33% during the first growing season post-disk, and -61% the second growing season. Confidence intervals of these effect sizes, however, included 0.
At the field level, there was no difference in Red-winged Blackbird density in buffer-bordered fields prior to disturbance (F2,11 = 1.50, P = 0.267). After disturbance had occurred, there was still no difference in blackbird density among the fields in 2008 (F2,11 = 1.42, P = 0.283) and 2009 (F2,11 = 0.58, P = 0.575). In 2008, relative effect sizes in blackbird density were 659% for burned fields and 108% for disked fields compared to control fields. In 2009, this difference was reduced to 17% for burned fields. At this time, the relative effect size between disked and control fields was -63%. Despite these large relative effect sizes, though, their respective confidence intervals included 0.
3.5. Indigo Buntings
Indigo Bunting density did not differ among agricultural conservation buffers prior to disturbance in 2007 (F4,38 = 1.12, P = 0.362). Based on relative effect sizes in 2008, bunting density in burned and disked buffers during their first growing season post-disturbance was greater than in control buffers (burned, 88%; disked, 118%). In 2009, bunting density in buffers during their first growing season post-burn was 12% greater relative to controls. Bunting density in disked buffers during their second growing season was 46% greater compared to controls. Despite these large effect sizes, however, there was no difference in bunting density regardless of type of or time since disturbance (2008, F2,36 = 3.18, P = 0.053; 2009, F4,41 = 1.49, P = 0.224).
Indigo Bunting density did not differ among buffer-bordered fields prior to disturbance in 2007 (F2,11 = 1.22, P = 0.331). Indigo Bunting density still did not differ among buffer-bordered fields after disturbance had occurred (2008, F2,11 = 0.77, P = 0.486; 2009, F2,11 = 1.38, P = 0.292). In 2008, relative effect sizes for bunting density between disturbed and control fields were -46% for burned fields and -17% for disked fields. These differences increased to -53% for burned fields and -23% for disked fields in 2009. Confidence intervals for these effects sizes, however, did include 0.
4. DISCUSSION
4.1. Avian Diversity and Density
Our results most frequently supported uniform detection functions, thus it is likely all birds present in the agricultural conservation buffers during transect surveys were detected. Similarly, based on a review of 75 published papers from 1985 to 2001 that conducted transect surveys of grassland birds, the probability of detection of Henslow’s (Ammodramus henslowii), Grasshopper, and Savannah Sparrows ranged from 0.93 to 1.0 when a bird was within 25 m of an observer [35Diefenbach DR, Brauning DW, Mattice JA. Variability in grassland bird counts related to observer differences and species detection rates. Auk 2003; 120: 1168-79.
[http://dx.doi.org/10.1642/0004-8038(2003)120[1168:VIGBCR]2.0.CO;2] ]. Due to uniform detection in the 0-15 m distance range, followed by an increase in detections in the 15-20 m distance range, avian behavior was likely not negatively influenced by observer presence and birds were attracted to the buffer edge where there was alternative vegetative cover (i.e., crop or non-crop habitat). If birds had been negatively influenced by observer presence, detections would have increased with increasing distance from the observer. Therefore, vegetation differences among buffers (e.g., vegetation height, vegetation density) and aversion behavior exhibited by birds in response to the observer did not influence the probability of detection during transect surveys.
Prescribed fire and light disking did not affect breeding bird diversity and density at either the buffer or field level during the first and second growing seasons post-disturbance. However, direction and magnitude of relative effect sizes on diversity and density for burned and disked buffers indicate the possibility of a type two error (i.e., failing to reject a false null hypothesis). If indeed our alternate hypothesis was true, this would be consistent with previous studies reporting differences in avian diversity and density between disturbed and undisturbed habitats. In northeast Nebraska, grassland bird abundance and diversity were greater in disked CRP fields than those that were unmanaged [36Negus LP, Davis CA, Wessel SE. Avian response to mid-contract management of conservation reserve program fields. Am Midl Nat 2010; 14: 296-310.
[http://dx.doi.org/10.1674/0003-0031-164.2.296] ]. In northwestern North Dakota, avian species richness and abundance of Bobolink, Grasshopper Sparrow, and Western Meadowlark were greater in burned than in unburned mixed-grass prairie sites [21Madden EM, Hansen AJ, Murphy RK. Influence of prescribed fire history on habitat and abundance of passerine birds in northern mixed-grass prairie. Can Field Nat 1999; 113: 627-40.]. These studies, however, occurred in large blocks of grassland habitat. Many grassland bird species, including Dickcissels, are area-sensitive species that prefer large habitat patches [37Bakker KK, Naugle DE, Higgins KF. Incorporating landscape attributes into models for migratory grassland bird conservation. Conserv Biol 2002; 16: 1638-46.
[http://dx.doi.org/10.1046/j.1523-1739.2002.01328.x] ]. Because agricultural conservation buffers only provide strip grassland habitat and cover a small area (0.17-3.49 ha in this study), disturbance methods used to maintain the early-successional state of buffers may not be an important factor in avian habitat selection at the field level. When applied in a rotational fashion, only ¼-⅓ of buffers is disturbed within any single year, leaving the remainder undisturbed, creating a mosaic of successional stages at the whole-field scale.
Although disturbance did not statistically affect diversity or density of breeding birds in our agricultural conservation buffers, disturbance does alter vegetation structure (e.g., vegetation height and density), which may influence a bird’s choice of habitat [38Best LB. The value of buffer habitats for birds in agricultural landscapes. In: Hohman WL, Halloum DJ, Eds. A comprehensive review of Farm Bill contributions to wildlife conservation, 1985-2000 Tech Rep 2000; 75-94.]. Burned mixed-grass prairies in north-central North Dakota had minimal vegetation coverage with little standing dead vegetation 1-year post-burn, but this increased and stabilized following the second growing season [39Grant TA, Madden E, Berkey GB. Tree and shrub invasion in northern-mixed-grass prairie: implications for breeding grassland birds. Wildl Soc Bull 2004; 32: 807-18.
[http://dx.doi.org/10.2193/0091-7648(2004)032[0807:TASIIN]2.0.CO;2] ]. In the agricultural conservation buffers at our study site, nest densities of Dickcissels and Red-winged Blackbirds were greater in burned buffers than in disked buffers [40Adams HL, Burger LW Jr, Riffell S. Disturbance and landscape effects on avian nests in agricultural conservation buffers. J Wildl Manage 2013; 77: 1213-20.
[http://dx.doi.org/10.1002/jwmg.568] ]. Alternatively, disking encourages the germination of forbs by disrupting grass-root structures and setting succession back further than burning [41Jones J, Coggin DS, Cummins JL, Hill J. Restoring and managing native prairies: A handbook for Mississippi landowners. Stoneville: Wildlife Mississippi 2007.]. In buffers used during this study, forb species richness in disked buffers was greater than in control and burned buffers during summer 2008 (P = 0.009-0.039) [42Dollar JG. Responses of butterfly and forb communities to management of semi-natural grassland buffers. Thesis 2011.]. In May 2009, these same buffers had a greater number of forb species than control and burned buffers (P = 0.009) [42Dollar JG. Responses of butterfly and forb communities to management of semi-natural grassland buffers. Thesis 2011.]. A greater presence of forbs in disked buffers can create suitable habitat for pollinating insects [17Dollar JG, Riffell SK, Burger LW Jr. Effects of managing semi-natural grassland buffers on butterflies. J Insect Conserv 2013; 7: 577-90.
[http://dx.doi.org/10.1007/s10841-012-9543-7] ], or other insect species that could serve as a food source for breeding grassland birds and their nestlings [43Mitchell KL, Riffell SK, Burger LW Jr, Vilella FJ. Nestling provisioning of Dickcissels in native warm-season grass field buffers. Wilson J Ornithol 2012; 124: 198-209.
[http://dx.doi.org/10.1676/11-152.1] ].
4.2. Dickcissels
Dickcissels are grassland specialists and a species of concern. From 1966 to 1979, Dickcissel populations in the Southeastern United States were declining at a rate of 6.1% every year [44Sauer JR, Hines JE, Fallon J. The North American breeding bird survey, results and analysis 1966-2007, version 515 2008. http://www. mbr-pwrc.usgs.gov/bbs/]. In recent years, however, Dickcissel populations have begun to increase. For instance, in the Mississippi Alluvial Valley; the physiographic region where our study site is located; the 2005-2015 Dickcissel population increased at a rate of 2.42% per year [45Sauer JR, Niven DK, Hines JE, et al. The North American breeding bird survey, results and analysis 1966–2015, version 207 2017. http:// www.mbr-pwrc.usgs.gov/bbs/]. Dickcissels likely do not require as large of early successional areas during the breeding season compared to other grassland birds (e.g., Eastern Meadowlark, Savannah Sparrow) [46Herkert JR. The effects of habitat fragmentation on Midwestern grassland bird communities. Ecol Appl 1994; 4: 461-71.
[http://dx.doi.org/10.2307/1941950] ]. Thus, Dickcissels may respond positively to conservation practices in fragmented areas, like those of the CRP in agricultural landscapes.
Dickcissels prefer grassland habitat with tall, dense vegetation structure [47Delisle JM, Savidge JA. Avian use and vegetation characteristics of conservation reserve program fields. J Wildl Manage 1997; 61: 318-25.
[http://dx.doi.org/10.2307/3802587] , 48Hughes JP, Robel RJ, Kemp KE, Zimmerman JL. Effects of habitat on Dickcissel abundance and nest success in conservation reserve program fields in Kansas. J Wildl Manage 1999; 63: 523-9.
[http://dx.doi.org/10.2307/3802638] ]. This may be one reason why Dickcissels were detected frequently in control buffers during this study. Because of the absence of disturbance, vegetation in these buffers provided many perching, singing, and foraging sites for birds. Disturbance temporarily alters this vegetation structure by reducing ground cover and vertical density. Time since disturbance can determine how quickly buffers can recover and once again accommodate Dickcissels during the breeding season. Dickcissels at our study site, for instance, had greater nest densities in burned buffers than those that were disked [40Adams HL, Burger LW Jr, Riffell S. Disturbance and landscape effects on avian nests in agricultural conservation buffers. J Wildl Manage 2013; 77: 1213-20.
[http://dx.doi.org/10.1002/jwmg.568] ].
Prescribed burning in agricultural conservation buffers did not significantly reduce Dickcissel density relative to the control buffer regardless of time since disturbance. Frequently, though, Dickcissel densities in grassland habitats decline immediately following a prescribed burn because of a lack of vegetative cover [23Fuhlendorf SD, Harrell WC, Engle DM, Hamilton RG, Davis CA, Leslie DM Jr. Should heterogeneity be the basis for conservation? Grassland bird response to fire and grazing. Ecol Appl 2006; 16(5): 1706-16.
[http://dx.doi.org/10.1890/1051-0761(2006)016[1706:SHBTBF]2.0.CO;2] [PMID: 17069365] ]. Given time, vegetation in burned buffers can recover quickly from disturbance because of warmer soil temperatures and greater availability of soil nutrients [49Harper CA, Bates GE, Hansbrough MP, Gudlin MJ, Gruchy JP, Keyser PD. Native warm-season grasses: Identification, establishment and management for wildlife and forage production in the mid-south. Knoxville: University of Tennessee Extension, PB1752 2007.]. This quick recovery may provide Dickcissels cover later in the breeding season. In central South Dakota, Dickcissel densities increased from early to late seral stages of a mixed-grass prairie [50Fritcher SC, Rumble MA, Flake LD. Grassland bird densities in seral stages of mixed-grass prairie. J Range Manage 2004; 57: 351-7.
[http://dx.doi.org/10.2307/4003858] ]. Additionally, Dickcissel abundance in Oklahoma grasslands increased with time since burning [18Olson DM, Wäckers FL. Management of field margins to maximize multiple ecological services. J Appl Ecol 2007; 44: 13-21.
[http://dx.doi.org/10.1111/j.1365-2664.2006.01241.x] ].
Alternatively, vegetation in disked buffers did not recover from disturbance as quickly as vegetation in burned buffers [41Jones J, Coggin DS, Cummins JL, Hill J. Restoring and managing native prairies: A handbook for Mississippi landowners. Stoneville: Wildlife Mississippi 2007.]. Because of this, Dickcissel density was greater in control and burned buffers than in disked buffers with respect to relative effect sizes. Though a greater abundance of forbs in disked buffers can provide Dickcissels foraging sites, these plants may not be able to provide optimal breeding and nesting sites for this species. Thus, Dickcissels occur in reduced abundance due to the lack of perching, singing, and nesting sites.
4.3. Red-winged Blackbirds
Red-winged Blackbirds, though commonly known as a wetland species, will breed in several habitat types throughout Mississippi, such as lowlands, weedy fields, roadways, and ditch banks [51Turcotte WH, Watts DL. Birds of Mississippi 1999.]. Blackbirds are also a facultative grassland species and will benefit from management practices used to promote grassland birds [52Patterson MP, Best LB. Bird abundance and nesting success in Iowa CRP fields: The importance of vegetation structure and composition. Am Midl Nat 1996; 135: 153-67.
[http://dx.doi.org/10.2307/2426881] , 53Vickery PD, Tubaro PL, Cardoso da Silva JM, Peterjohn BG, Herkert JR, Cavalcanti RB. Conservation of grassland birds in the western hemisphere. Stud Avian Biol 1999; 19: 2-26.]. Furthermore, this species is known commonly as a crop pest and their presence in agricultural conservation buffers may be viewed as a drawback to this conservation practice by land managers. For instance, the flocking and foraging behavior of Red-winged Blackbirds in the northern Great Plains causes extensive damage to sunflower (Helianthus annuus) [54Linz GM, Klug PE, Dolbeer RA. Ecology and management of red-winged blackbirds. In: Linz GM, Avery ML, Dolbeer RA, Eds. Ecology and management of blackbirds (Icteridae) in North America 2017; 17-41.
[http://dx.doi.org/10.4324/9781315156439-2] ]. However, in the southeastern United States, cultivated rice may make up a large portion of the birds’ diet [54Linz GM, Klug PE, Dolbeer RA. Ecology and management of red-winged blackbirds. In: Linz GM, Avery ML, Dolbeer RA, Eds. Ecology and management of blackbirds (Icteridae) in North America 2017; 17-41.
[http://dx.doi.org/10.4324/9781315156439-2] ], which is a crop not typically grown in upland areas where buffers are more commonly established.
Red-winged Blackbird density in burned buffers did not differ from control buffers in 2008 and 2009. In a Kansas study, Red-winged Blackbird density was greater in unburned than burned sites, but this difference was not significant [55Robel RJ, Hughes JP, Hull SD, Kemp KE, Klute DS. Spring burning: resulting avian abundance and nesting in Kansas CRP. J Range Manage 1998; 51: 132-8.
[http://dx.doi.org/10.2307/4003197] ]. Red-winged Blackbird densities in an east-central North Dakota mixed-grass prairie increased 2 to 5 years post-burn [28Johnson DH. Effects of fire on bird populations in mixed-grass prairie. In: Knopf FL, Samson FB, Eds. Ecology and conservation of Great Plains vertebrates 1997; 181-206.
[http://dx.doi.org/10.1007/978-1-4757-2703-6_8] ]. Though there was no significant difference in blackbird densities between burned and unburned buffers in this study, we did not collect data past the second growing season post-disturbance. Thus, it is possible a difference in blackbird density between these 2 disturbance types may have been detected had the study continued during subsequent years [28Johnson DH. Effects of fire on bird populations in mixed-grass prairie. In: Knopf FL, Samson FB, Eds. Ecology and conservation of Great Plains vertebrates 1997; 181-206.
[http://dx.doi.org/10.1007/978-1-4757-2703-6_8] ].
Though there were no significant differences found in this study, Red-winged Blackbird density in 2008 was greater in control buffers than in disked buffers based on relative effect sizes. In 2009, this difference was still existent in these same buffers during their second growing season after disking. Like Dickcissels, Red-winged Blackbirds are associated positively with vertical vegetation density [52Patterson MP, Best LB. Bird abundance and nesting success in Iowa CRP fields: The importance of vegetation structure and composition. Am Midl Nat 1996; 135: 153-67.
[http://dx.doi.org/10.2307/2426881] ]. Thus, these birds may avoid disked buffers because of a lack of foraging and breeding habitat.
Red-winged blackbirds are known for causing excessive crop damage. In North Dakota, South Dakota, and Nebraska, for instance, the average annual impacts of Red-winged Blackbirds, along with Yellow-headed Blackbirds (Xanthocephalus xanthocephalus) and Common Grackles (Quiscalus quiscula), on sunflower production from 2009 to 2013 were US $18.7 million, US $7.3 million, and US $2.6 million, respectively [56Ernst K, Elser J, Linz G, et al. The economic impacts of blackbird (Icteridae) damage to sunflower in the USA. Pest Manag Sci 2019; 75(11): 2910-5.
[http://dx.doi.org/10.1002/ps.5486] [PMID: 31087456] ]. In regards to our study, planted crops included soybean, corn, and Bermudagrass. We did not observe any occurrences of crop damage by Red-winged Blackbirds to these crops while performing transect surveys. Furthermore, in a concurrent study, we determined Red-winged Blackbird nest success was only 12.9% at the peak of their breeding season (early August), thus buffers may serve only as sink habitat for this species [40Adams HL, Burger LW Jr, Riffell S. Disturbance and landscape effects on avian nests in agricultural conservation buffers. J Wildl Manage 2013; 77: 1213-20.
[http://dx.doi.org/10.1002/jwmg.568] ]. However, land managers should be attentive to potential crop damage by Red-winged Blackbirds or similar species.
4.4. Indigo Buntings
Indigo Buntings are a common forest-field edge species that will utilize woodland and grassland habitat in close proximity [57Weldon AJ, Haddad NM. The effects of patch shape on Indigo Buntings: Evidence for an ecological trap. Ecology 2005; 86: 1422-31.
[http://dx.doi.org/10.1890/04-0913] ]. These birds may also be found in shrubby areas and weedy fields [58Payne RB. Indigo bunting (Passerina cyanea), version 2.0. In: Poole AF, Ed. Birds of North America 2006.]. Indigo Buntings may also prefer edge habitat more irregular in shape than linear habitat [57Weldon AJ, Haddad NM. The effects of patch shape on Indigo Buntings: Evidence for an ecological trap. Ecology 2005; 86: 1422-31.
[http://dx.doi.org/10.1890/04-0913] , 59Brennan SP, Schnell GD. Relationship between bird abundances and landscape characteristics: The influence of scale. Environ Monit Assess 2005; 105(1-3): 209-28.
[http://dx.doi.org/10.1007/s10661-005-3694-x] [PMID: 15952521] ], such as that commonly provided by agricultural conservation buffers.
As with Dickcissels and Red-winged Blackbirds, Indigo Bunting density did not vary significantly between disturbed and undisturbed buffers during this study. Several studies, however, have shown Indigo Buntings respond positively to disturbance. In North American oak (Quercus spp.) savannahs, for instance, bunting densities increased following prescribed burns [60Brawn JD, Robinson SK, Thompson FR III. The role of disturbance in the ecology and conservation of birds. Annu Rev Ecol Syst 2001; 32: 251-76.
[http://dx.doi.org/10.1146/annurev.ecolsys.32.081501.114031] ]. Indigo Buntings have also been frequently observed in New Hampshire clear-cut forest sites [61Costello CA, Yamasaki M, Pekins PJ, Leak WB, Neefus CD. Songbirds response to group selection harvests and clearcuts in a New Hampshire northern hardwood forest. For Ecol Manage 2000; 127: 41-54.
[http://dx.doi.org/10.1016/S0378-1127(99)00131-0] ]. However, Indigo Bunting density can be least immediately following disturbance due to lack of vegetative cover. Frequent burning, for instance, can reduce shrub density and, in turn, Indigo Bunting density [62Brennan LA, Engstrom RT, Palmer WE, et al. Whither wildlife without fire? Trans North Am Wildl Nat Resour Conf 1998; 63: 402-14.]. Also, Indigo Buntings preferred areas with dense herbaceous ground cover when white-tailed deer (Odocoileus virginianus) were excluded from forested sites in Virginia-Indigo Bunting density increased with time since exclusion [63McShea WJ, Rappole JH. Managing the abundance and diversity of breeding bird populations through manipulation of deer populations. Conserv Biol 2000; 14: 1161-70.
[http://dx.doi.org/10.1046/j.1523-1739.2000.99210.x] ].
Unmanaged agricultural conservation buffers may be able to provide more cover for Indigo Buntings than those that have recently been disturbed. With time, however, disturbed buffers could be preferred Indigo Bunting habitat as vegetation density in unmanaged buffers increases.
CONCLUSION
We were unable to support our hypothesis that prescribed fire is the best form of periodic disturbance to manage agricultural conservation buffers for breeding grassland birds. Though disturbance did not significantly affect diversity or density of breeding birds in the buffers, relative effect sizes indicated a potential biological significance in avian diversity and density between burned and disked buffers, possibly promoting the use of burning over disking.
Fire effects on herbaceous vegetation in agricultural conservation buffers will differ with frequency and intensity of burns, amount and type of litter cover, soil and biomass moisture, temperature, and wind speed [47Delisle JM, Savidge JA. Avian use and vegetation characteristics of conservation reserve program fields. J Wildl Manage 1997; 61: 318-25.
[http://dx.doi.org/10.2307/3802587] ]. This can benefit other vertebrate species other than birds, such as amphibians, reptiles, and small mammals [62Brennan LA, Engstrom RT, Palmer WE, et al. Whither wildlife without fire? Trans North Am Wildl Nat Resour Conf 1998; 63: 402-14.]. Given variation among these aforementioned factors with both time and space, using prescribed burning can also lead to a heterogeneous landscape [64Parr CL, Andersen AN. Patch mosaic burning for biodiversity conservation: A critique of the pyrodiversity paradigm. Conserv Biol 2006; 20(6): 1610-9.
[http://dx.doi.org/10.1111/j.1523-1739.2006.00492.x] [PMID: 17181796] ]. In Australia, for instance, the use of a patch mosaic burn (PMB) may further encourage landscape heterogeneity and biodiversity [64Parr CL, Andersen AN. Patch mosaic burning for biodiversity conservation: A critique of the pyrodiversity paradigm. Conserv Biol 2006; 20(6): 1610-9.
[http://dx.doi.org/10.1111/j.1523-1739.2006.00492.x] [PMID: 17181796] ]. This technique, however, was not feasible given our buffers occurred in small, linear strips ranging from 0.17 to 3.12 ha, greatly limiting the number of ignition points [65Parr CL, Brockett BH. Patch-mosaic burning: A new paradigm for savanna fire management in protected areas? Koedoe 1999; 42: 117-30.
[http://dx.doi.org/10.4102/koedoe.v42i2.237] ]. The buffers in our study area, however, existed in a heterogeneous landscape that included pastures, woodlands, wetlands, and ponds. Such varying types of land cover may contribute to increasing biodiversity of flora and fauna in and around the buffers.
Prescribed burning for this study was performed during the dormant season in mid- to late March. This time is beneficial because it briefly removes winter cover for wildlife and it reduces the likelihood of harming breeding birds and their young. There is a risk, though, of negatively influencing other populations inhabiting the buffers, specifically insects that serve as an important food source for breeding birds. For instance, the abundance of litter-dwelling insects decreased after a prescribed burn in small prairies of the Midwestern United States [66Panzer R. Compatibility of prescribed burning with the conservation of insects in small, isolated prairie reserves. Conserv Biol 2002; 16: 1296-307.
[http://dx.doi.org/10.1046/j.1523-1739.2002.01077.x] ]. To maintain agricultural conservation buffers as early-successional habitat for Northern Bobwhite (as was required in the buffers of our study), a disturbance is necessary to prevent woody plant encroachment. However, the inclusion of forbs, such as black-eyed susan and partridge pea in our buffers, will encourage greater insect populations in the buffers, particularly pollinating insects.
Disking can be used as an alternative to fire when the landscape does not permit safe burns. Disking can encourage biomass decomposition, expose bare ground, and increase nutrient availability [49Harper CA, Bates GE, Hansbrough MP, Gudlin MJ, Gruchy JP, Keyser PD. Native warm-season grasses: Identification, establishment and management for wildlife and forage production in the mid-south. Knoxville: University of Tennessee Extension, PB1752 2007.]. Additionally, disking can promote insect activity by setting further back than burning, allowing forbs to become the dominant vegetation group. In the same buffers as our study, butterfly abundance increased 1- to 2-years post-disking due to the increase in forb abundance in the buffers [17Dollar JG, Riffell SK, Burger LW Jr. Effects of managing semi-natural grassland buffers on butterflies. J Insect Conserv 2013; 7: 577-90.
[http://dx.doi.org/10.1007/s10841-012-9543-7] ]. Thus, we strongly recommend landowners consider their objectives when establishing and managing agricultural conservation buffers around their crop production fields.
ETHICS APPROVAL AND CONSENT TO PARTICIPATE
Not applicable.
HUMAN AND ANIMAL RIGHTS
No animals/humans were used for studies that are the basis of this research.
CONSENT FOR PUBLICATION
Not applicable.
AVAILABILITY OF DATA AND MATERIAL
Data supporting the findings of the article are available in the Mississippi State University Institutional Repository at https://hdl.handle.net/11668/16484.
FUNDING
This study was funded by the United States Department of Agriculture (USDA) Natural Resources Conservation Service (NRCS) grant number 69-7482-6-286. Additional financial support was provided by the Mississippi Agricultural and Forestry Experiment Station (MAFES), the Mississippi State University Forest and Wildlife Research Center, and the Mississippi State University College of Forest Resources.
CONFLICT OF INTEREST
The authors declare no conflict of interest, financial or otherwise.
ACKNOWLEDGEMENTS
Special thanks to J. Dollar, K. Mitchell, S. Hale, K. Allender, R. Hicks, A. Carnes, R. Bradford, A. Hollander, and J. Nunley for their assistance with data collection, and J. Adams for editorial assistance. This paper is dedicated to the memory of our co-author and friend, Dr. Sam Riffell, who was instrumental in the planning and implementation of this study.
REFERENCES
| [1] | Smeding FW, Joenje W. Farm-nature plan: Landscape ecology based on farm planning. Landsc Urban Plan 1999; 46: 109-15. [http://dx.doi.org/10.1016/S0169-2046(99)00052-3] |
| [2] | Marshall EJP, Moonen AC. Field margins in northern Europe: Their functions and interactions with agriculture. Agric Ecosyst Environ 2002; 89: 5-21. [http://dx.doi.org/10.1016/S0167-8809(01)00315-2] |
| [3] | Liira J, Aavik T, Parrest O, Zobel M. Agricultural sector, rural environment and biodiversity in the central and eastern European EU member states. AGD Landsc Environ 2008; 2: 46-64. |
| [4] | Robinson RA, Sutherland WJ. Post-war changes in arable farming and biodiversity in Great Britain. J Appl Ecol 2002; 39: 157-76. [http://dx.doi.org/10.1046/j.1365-2664.2002.00695.x] |
| [5] | United States Department of Agriculture. National Agricultural Statistics Service [accessed 2018 May 22] Available from: https://www.nass.usda.gov/ |
| [6] | Benton TG, Vickery JA, Wilson JD. Farmland biodiversity: Is habitat heterogeneity the key? Trends Ecol Evol 2003; 18: 182-8. [http://dx.doi.org/10.1016/S0169-5347(03)00011-9] |
| [7] | Burger LW Jr, Smith MD, Barbour PJ, Hamrick R. Grassland bird response to agricultural field borders. Forest Wildl Res Center Res Adv 2005; 8: 1-4. |
| [8] | Green RE, Cornell SJ, Scharlemann JPW, Balmford A. Farming and the fate of wild nature. Science 2005; 307(5709): 550-5. [http://dx.doi.org/10.1126/science.1106049] [PMID: 15618485] |
| [9] | Samson F, Knopf K. Prairie conservation in North America. Bioscience 1994; 44: 418-21. [http://dx.doi.org/10.2307/1312365] |
| [10] | Knopf FL. Avian assemblages on altered grasslands. Stud Avian Biol 1994; 15: 247-57. |
| [11] | Donal PF, Gree RE, Heath MF. Agricultural intensification and the collapse of Europe’s farmland bird populations. Proc Biol Sci 2001; 268(1462): 25-9. [http://dx.doi.org/10.1098/rspb.2000.1325] [PMID: 12123294] |
| [12] | Pedroli GBM, Meiner A. Landscapes in transition: An account of 25 years of land cover change in Europe. Publications Office of the European Union, EEA Report No 10/2017 2017. |
| [13] | Barbour PJ, Martin SW, Burger W. Estimating economic impact of conservation field borders on farm revenue. Crop Manag 2007; 6 [http://dx.doi.org/10.1094/CM-2007-0614-01-RS] |
| [14] | Josefsson J, Berg A, Hiron M, Pärt T, Eggers S. Grass buffer strips benefit invertebrate and breeding skylark numbers in a heterogeneous agricultural landscape. Agric Ecosyst Environ 2013; 181: 101-7. [http://dx.doi.org/10.1016/j.agee.2013.09.018] |
| [15] | Evans KO, Burger LW Jr, Riffell S, Smith MD. Assessing multiregion avian benefits from strategically targeted agricultural buffers. Conserv Biol 2014; 28(4): 892-901. [http://dx.doi.org/10.1111/cobi.12311] [PMID: 24779636] |
| [16] | Lovell ST, Sullivan WC. Environmental benefits of conservation buffers in the United States: evidence, promise, and open questions. Agric Ecosyst Environ 2006; 112: 249-60. [http://dx.doi.org/10.1016/j.agee.2005.08.002] |
| [17] | Dollar JG, Riffell SK, Burger LW Jr. Effects of managing semi-natural grassland buffers on butterflies. J Insect Conserv 2013; 7: 577-90. [http://dx.doi.org/10.1007/s10841-012-9543-7] |
| [18] | Olson DM, Wäckers FL. Management of field margins to maximize multiple ecological services. J Appl Ecol 2007; 44: 13-21. [http://dx.doi.org/10.1111/j.1365-2664.2006.01241.x] |
| [19] | McCoy TD, Kurzejeski EW, Burger LW Jr, Ryan MR. Effects of conservation practice, mowing, and temporal changes on vegetation structure on Conservation Reserve Program fields in northern Missouri. Wildl Soc Bull 2001; 29: 979-87. |
| [20] | Rodenhouse NL, Best LB, O’Connor RJ, Bollinger EK. Effects of agricultural practices on farmland structures. In: Martin TE, Finch DM, Eds. Ecology and management of Neotropical migratory birds: A synthesis and review of critical issues 1995; 269-93. |
| [21] | Madden EM, Hansen AJ, Murphy RK. Influence of prescribed fire history on habitat and abundance of passerine birds in northern mixed-grass prairie. Can Field Nat 1999; 113: 627-40. |
| [22] | Reinking DL. Fire regimes and avian responses in the central tallgrass prairie. Stud Avian Biol 2005; 30: 116-26. |
| [23] | Fuhlendorf SD, Harrell WC, Engle DM, Hamilton RG, Davis CA, Leslie DM Jr. Should heterogeneity be the basis for conservation? Grassland bird response to fire and grazing. Ecol Appl 2006; 16(5): 1706-16. [http://dx.doi.org/10.1890/1051-0761(2006)016[1706:SHBTBF]2.0.CO;2] [PMID: 17069365] |
| [24] | Douglas DJT, Vickery JA, Benton TG. Improving the value of field margins as foraging habitat for farmland birds. J Appl Ecol 2009; 46: 353-62. [http://dx.doi.org/10.1111/j.1365-2664.2009.01613.x] |
| [25] | Murphy RK, Shaffer TL, Grant TA, Derrig JL, Rubin CS, Kerns CK. Sparrow nest survival in relation to prescribed fire and woody plant invasion in a northern mixed-grass prairie. Wildl Soc Bull 2017; 41: 442-52. [http://dx.doi.org/10.1002/wsb.780] |
| [26] | Engstrom RT, Vickery PD, Perkins DW, Shriver WG. Effects of fire regime on birds in southeastern pine savannas and native prairies. Stud Avian Biol 2005; 30: 147-60. |
| [27] | Haikerwal A, Reisen F, Sim MR, et al. Impact of smoke from prescribed burning: Is it a public health concern? J Air Waste Manag Assoc 2015; 65(5): 592-8. [http://dx.doi.org/10.1080/10962247.2015.1032445] [PMID: 25947317] |
| [28] | Johnson DH. Effects of fire on bird populations in mixed-grass prairie. In: Knopf FL, Samson FB, Eds. Ecology and conservation of Great Plains vertebrates 1997; 181-206. [http://dx.doi.org/10.1007/978-1-4757-2703-6_8] |
| [29] | Burger LW Jr, McKenzie D, Thackston R, Demaso SJ. The role of farm policy in achieving large-scale conservation: Bobwhite and buffers. Wildl Soc Bull 2006; 34: 986-93. [http://dx.doi.org/10.2193/0091-7648(2006)34[986:TROFPI]2.0.CO;2] |
| [30] | Buckland ST, Anderson DR, Burnham KP, Laake JL, Borchers DL, Thomas L. Introduction to distance sampling 2001. |
| [31] | Smith MD, Barbour PJ, Burger LW Jr, Dinsmore SJ. Breeding bird abundance and diversity in agricultural field borders in the Black Belt Prairie of Mississippi. Proc Annu Conf Southeast Assoc Fish Wildl Agencies 2005; 43-56. |
| [32] | Shannon CE, Weaver W. The mathematical theory of communication 1963. |
| [33] | SAS 9.4 product documentation. 2013. |
| [34] | Thomas L, Buckland ST, Rexstad EA, et al. Distance software: design and analysis of distance sampling surveys for estimating population size. J Appl Ecol 2010; 47(1): 5-14. [http://dx.doi.org/10.1111/j.1365-2664.2009.01737.x] [PMID: 20383262] |
| [35] | Diefenbach DR, Brauning DW, Mattice JA. Variability in grassland bird counts related to observer differences and species detection rates. Auk 2003; 120: 1168-79. [http://dx.doi.org/10.1642/0004-8038(2003)120[1168:VIGBCR]2.0.CO;2] |
| [36] | Negus LP, Davis CA, Wessel SE. Avian response to mid-contract management of conservation reserve program fields. Am Midl Nat 2010; 14: 296-310. [http://dx.doi.org/10.1674/0003-0031-164.2.296] |
| [37] | Bakker KK, Naugle DE, Higgins KF. Incorporating landscape attributes into models for migratory grassland bird conservation. Conserv Biol 2002; 16: 1638-46. [http://dx.doi.org/10.1046/j.1523-1739.2002.01328.x] |
| [38] | Best LB. The value of buffer habitats for birds in agricultural landscapes. In: Hohman WL, Halloum DJ, Eds. A comprehensive review of Farm Bill contributions to wildlife conservation, 1985-2000 Tech Rep 2000; 75-94. |
| [39] | Grant TA, Madden E, Berkey GB. Tree and shrub invasion in northern-mixed-grass prairie: implications for breeding grassland birds. Wildl Soc Bull 2004; 32: 807-18. [http://dx.doi.org/10.2193/0091-7648(2004)032[0807:TASIIN]2.0.CO;2] |
| [40] | Adams HL, Burger LW Jr, Riffell S. Disturbance and landscape effects on avian nests in agricultural conservation buffers. J Wildl Manage 2013; 77: 1213-20. [http://dx.doi.org/10.1002/jwmg.568] |
| [41] | Jones J, Coggin DS, Cummins JL, Hill J. Restoring and managing native prairies: A handbook for Mississippi landowners. Stoneville: Wildlife Mississippi 2007. |
| [42] | Dollar JG. Responses of butterfly and forb communities to management of semi-natural grassland buffers. Thesis 2011. |
| [43] | Mitchell KL, Riffell SK, Burger LW Jr, Vilella FJ. Nestling provisioning of Dickcissels in native warm-season grass field buffers. Wilson J Ornithol 2012; 124: 198-209. [http://dx.doi.org/10.1676/11-152.1] |
| [44] | Sauer JR, Hines JE, Fallon J. The North American breeding bird survey, results and analysis 1966-2007, version 515 2008. http://www. mbr-pwrc.usgs.gov/bbs/ |
| [45] | Sauer JR, Niven DK, Hines JE, et al. The North American breeding bird survey, results and analysis 1966–2015, version 207 2017. http:// www.mbr-pwrc.usgs.gov/bbs/ |
| [46] | Herkert JR. The effects of habitat fragmentation on Midwestern grassland bird communities. Ecol Appl 1994; 4: 461-71. [http://dx.doi.org/10.2307/1941950] |
| [47] | Delisle JM, Savidge JA. Avian use and vegetation characteristics of conservation reserve program fields. J Wildl Manage 1997; 61: 318-25. [http://dx.doi.org/10.2307/3802587] |
| [48] | Hughes JP, Robel RJ, Kemp KE, Zimmerman JL. Effects of habitat on Dickcissel abundance and nest success in conservation reserve program fields in Kansas. J Wildl Manage 1999; 63: 523-9. [http://dx.doi.org/10.2307/3802638] |
| [49] | Harper CA, Bates GE, Hansbrough MP, Gudlin MJ, Gruchy JP, Keyser PD. Native warm-season grasses: Identification, establishment and management for wildlife and forage production in the mid-south. Knoxville: University of Tennessee Extension, PB1752 2007. |
| [50] | Fritcher SC, Rumble MA, Flake LD. Grassland bird densities in seral stages of mixed-grass prairie. J Range Manage 2004; 57: 351-7. [http://dx.doi.org/10.2307/4003858] |
| [51] | Turcotte WH, Watts DL. Birds of Mississippi 1999. |
| [52] | Patterson MP, Best LB. Bird abundance and nesting success in Iowa CRP fields: The importance of vegetation structure and composition. Am Midl Nat 1996; 135: 153-67. [http://dx.doi.org/10.2307/2426881] |
| [53] | Vickery PD, Tubaro PL, Cardoso da Silva JM, Peterjohn BG, Herkert JR, Cavalcanti RB. Conservation of grassland birds in the western hemisphere. Stud Avian Biol 1999; 19: 2-26. |
| [54] | Linz GM, Klug PE, Dolbeer RA. Ecology and management of red-winged blackbirds. In: Linz GM, Avery ML, Dolbeer RA, Eds. Ecology and management of blackbirds (Icteridae) in North America 2017; 17-41. [http://dx.doi.org/10.4324/9781315156439-2] |
| [55] | Robel RJ, Hughes JP, Hull SD, Kemp KE, Klute DS. Spring burning: resulting avian abundance and nesting in Kansas CRP. J Range Manage 1998; 51: 132-8. [http://dx.doi.org/10.2307/4003197] |
| [56] | Ernst K, Elser J, Linz G, et al. The economic impacts of blackbird (Icteridae) damage to sunflower in the USA. Pest Manag Sci 2019; 75(11): 2910-5. [http://dx.doi.org/10.1002/ps.5486] [PMID: 31087456] |
| [57] | Weldon AJ, Haddad NM. The effects of patch shape on Indigo Buntings: Evidence for an ecological trap. Ecology 2005; 86: 1422-31. [http://dx.doi.org/10.1890/04-0913] |
| [58] | Payne RB. Indigo bunting (Passerina cyanea), version 2.0. In: Poole AF, Ed. Birds of North America 2006. |
| [59] | Brennan SP, Schnell GD. Relationship between bird abundances and landscape characteristics: The influence of scale. Environ Monit Assess 2005; 105(1-3): 209-28. [http://dx.doi.org/10.1007/s10661-005-3694-x] [PMID: 15952521] |
| [60] | Brawn JD, Robinson SK, Thompson FR III. The role of disturbance in the ecology and conservation of birds. Annu Rev Ecol Syst 2001; 32: 251-76. [http://dx.doi.org/10.1146/annurev.ecolsys.32.081501.114031] |
| [61] | Costello CA, Yamasaki M, Pekins PJ, Leak WB, Neefus CD. Songbirds response to group selection harvests and clearcuts in a New Hampshire northern hardwood forest. For Ecol Manage 2000; 127: 41-54. [http://dx.doi.org/10.1016/S0378-1127(99)00131-0] |
| [62] | Brennan LA, Engstrom RT, Palmer WE, et al. Whither wildlife without fire? Trans North Am Wildl Nat Resour Conf 1998; 63: 402-14. |
| [63] | McShea WJ, Rappole JH. Managing the abundance and diversity of breeding bird populations through manipulation of deer populations. Conserv Biol 2000; 14: 1161-70. [http://dx.doi.org/10.1046/j.1523-1739.2000.99210.x] |
| [64] | Parr CL, Andersen AN. Patch mosaic burning for biodiversity conservation: A critique of the pyrodiversity paradigm. Conserv Biol 2006; 20(6): 1610-9. [http://dx.doi.org/10.1111/j.1523-1739.2006.00492.x] [PMID: 17181796] |
| [65] | Parr CL, Brockett BH. Patch-mosaic burning: A new paradigm for savanna fire management in protected areas? Koedoe 1999; 42: 117-30. [http://dx.doi.org/10.4102/koedoe.v42i2.237] |
| [66] | Panzer R. Compatibility of prescribed burning with the conservation of insects in small, isolated prairie reserves. Conserv Biol 2002; 16: 1296-307. [http://dx.doi.org/10.1046/j.1523-1739.2002.01077.x] |
| [67] | Hamel PB. Land manager’s guide to the birds of the South 1992. [http://dx.doi.org/10.2737/SE-GTR-22] |