- Home
- About Journals
-
Information for Authors/ReviewersEditorial Policies
Publication Fee
Publication Cycle - Process Flowchart
Online Manuscript Submission and Tracking System
Publishing Ethics and Rectitude
Authorship
Author Benefits
Reviewer Guidelines
Guest Editor Guidelines
Peer Review Workflow
Quick Track Option
Copyediting Services
Bentham Open Membership
Bentham Open Advisory Board
Archiving Policies
Fabricating and Stating False Information
Post Publication Discussions and Corrections
Editorial Management
Advertise With Us
Funding Agencies
Rate List
Kudos
General FAQs
Special Fee Waivers and Discounts
- Contact
- Help
- About Us
- Search


The Open Paleontology Journal
(Discontinued)
ISSN: 1874-4257 ― Volume 5, 2014
Pleistocene Planktic Foraminiferal Events in the Northwest Pacific Near Japan
Hanako Domitsu*, Motoyoshi Oda
Abstract
Using an age model based on magnetostratigraphic and tephrostratigraphic control points, we assigned ages to two planktic foraminiferal events from the Pleistocene sequence in the northwest Pacific: the first occurrence of Neogloboquadrina pachyderma at 1.25 ± 0.04 Ma and the last occurrence of Neogloboquadrina inglei at 0.73 ± 0.05 Ma.
Article Information
Identifiers and Pagination:
Year: 2008Volume: 1
First Page: 1
Last Page: 6
Publisher Id: TOPALOJ-1-1
DOI: 10.2174/1874425700801010001
Article History:
Received Date: 11/12/2007Revision Received Date: 1/2/2008
Acceptance Date: 1/2/2008
Electronic publication date: 11/3/2008
Collection year: 2008
open-access license: This is an open access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.5/), which permits unrestrictive use, distribution, and reproduction in any medium, provided the original work is properly cited.
* Address correspondence to this author at the Institute of Geology and Paleontology, Graduate School of Science, Tohoku University, 6-3 Aoba, Aramaki, Aoba-ku, Sendai 980-8578, Japan; Fax: +81-22-795-6634; E-mail: hana@mail.tains.tohoku.ac.jp
| Open Peer Review Details | |||
|---|---|---|---|
| Manuscript submitted on 11-12-2007 |
Original Manuscript | Pleistocene Planktic Foraminiferal Events in the Northwest Pacific Near Japan | |
INTRODUCTION
Planktic foraminiferal biostratigraphy is a standard tool for the global correlation of marine strata, especially in tropical to subtropical regions. At mid to high latitudes, a rather small number of temperate to polar species need to be applied to sequencing the Pleistocene through bioevents and biozones. The Neogloboquadrina group is representative of the planktic foraminifera typical of late Neogene mid- to high-latitude regions. Kucera and Kennett [1] reported the biochronology of eight neogloboquadrinid events (first and last occurrences) in six Pliocene to Pleistocene deep-sea sequences obtained during Ocean Drilling Program (ODP) Leg 167 in the northeast Pacific near California. They described a new species, Neogloboquadrina inglei, the last occurrence (LO) of which was ca. 0.71 Ma in these sequences [1].
During ODP Leg 186, Sites 1150 and 1151 were drilled on the deep-sea terrace along the Japan Trench in the northwest Pacific. Motoyama et al. [2] established age models for Sites 1150 and 1151 based on a combination of magnetostratigraphy, tephrostratigraphy, and siliceous and calcareous microfossil biostratigraphy, and proposed a regional late Cenozoic magnetobiochronological timescale for northern Japan and its adjacent areas. To date, no planktic foraminiferal events have been recognized in the Pleistocene after the LO of Neogloboquadrina asanoi at ca. 1.8 Ma [2]. In the present study, we present two new biostratigraphic events, to improve the planktic foraminiferal biostratigraphy of the uppermost Pliocene and Pleistocene of the northwestern Pacific (Site 1150).
STUDY SITE AND SAMPLES
The studied ODP Hole 1150A is located at 39˚11'N, 143˚20'E, 2681 m in water depth (Fig. 1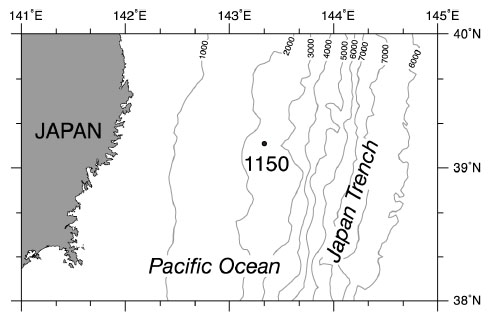 ). The age model for Site 1150 indicates a relatively constant sedimentation rate of 6 cm/kyr during the Late Pliocene and Pleistocene (3–0.09 Ma) and a high sedimentation rate of 30 cm/kyr during the past 90 kyr [2].
). The age model for Site 1150 indicates a relatively constant sedimentation rate of 6 cm/kyr during the Late Pliocene and Pleistocene (3–0.09 Ma) and a high sedimentation rate of 30 cm/kyr during the past 90 kyr [2].
 |
Fig. (1) Location of Site 1150 near the Japan Trench in the northwest Pacific drilled during ODP Leg 186. |
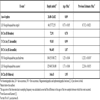
We examined a total of 100 samples above the base of Subchron C2n (Olduvai) from the upper 140 m of the core. Details of the sample preparation techniques have been described previously [3]. We selected all specimens from representative aliquots of individuals >125 µm from each sample. We calculated the ages of bioevents by linear interpolation between the two nearest magnetostratigraphic or tephrostratigraphic control points (Table 1).
RESULTS AND DISCUSSION
At Site 1150, the sedimentary sequence above the Aso-4 tephra (ca. 0.09 Ma [5, 6]) at 26.8 m below the seafloor (mbsf) contained generally abundant and well preserved planktic foraminifera. Below the tephra planktic foraminifera were sporadically rare or absent. Consequently, we established the planktic foraminiferal biostratigraphy based on 24 horizons from the upper 140 m of Hole 1150A (Fig. 2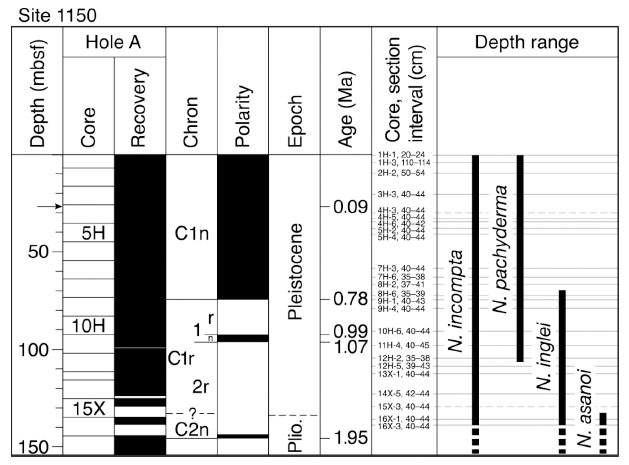 ). The average time resolution is 70–110 kyr below the Aso-4 tephra and 23 kyr above the tephra.
). The average time resolution is 70–110 kyr below the Aso-4 tephra and 23 kyr above the tephra.

After the LO of N. asanoi between 129.4 and 136 mbsf, two neogloboquadrinid events, the first occurrence (FO) of Neogloboquadrina pachyderma and the LO of N. inglei, were observed in the Pleistocene sequence at Site 1150 (Table 1). We recognized the FO of N. pachyderma between 104.55 and 108.72 mbsf, and this species occurs continuously throughout the rest of the Pleistocene (Fig. 2). In the present study, N. pachyderma exhibits both sinistral and dextral coiling, but dextrally coiled forms represent less than 3% of the sample. The ratio of dextrally coiled forms to the total number of N. pachyderma at Site 1150 is consistent with that in surface sediments around Japan [Oda M, unpublished data, 1983]. The N. pachyderma from this study appears to correspond to N. pachyderma B from the California margin reported by Kucera and Kennett [1]. According to our age model, N. pachyderma first occurred 1.25 ± 0.04 Ma at Site 1150 (Table 1). This age is concordant with the mean age of 1.23 ± 0.02 Ma for the FO of N. pachyderma B in the California margin [1].
We detected the LO of N. inglei between 66.57 and 72.55 mbsf, just above the base of Chron C1n (Brunhes; Fig. 2), and estimated the age to be 0.73 ± 0.05 Ma (Table 1). This estimate is similar to the mean age of 0.71 ± 0.02 Ma for the LO of N. inglei in the California margin [1], suggesting that the LO of N. inglei is a synchronous event between the eastern and western North Pacific. These two planktic foraminiferal events are useful biochronological markers for northern Japan and mid- to high-latitude regions in the North Pacific.
TAXONOMIC NOTES
The following comments briefly summarize principal morphologic characteristics of the four species presented in this study. All are illustrated by scanning electron micrographs (Plates 1 -3
-3 ).
).

Neogloboquadrina pachyderma (Ehrenberg) (Plate 1, Fig. 1)
Aristerospira pachyderma Ehrenberg, 1861, p. 276, 277, 303 [7]; Ehrenberg, 1873, (Plate 1, Fig. 4) [8].
Test small, dominantly sinistrally coiled, rounded in both axial and equatorial peripheries. Four globular chambers in the final whorl, rapidly increasing in size as added, the final chamber frequently forming a bulla-like cover over much of the umbilicus. Aperture umbilical, interiomarginal, a low arch with a distinct rim-like lip.
Neogloboquadrina inglei Kucera and Kennett (Plate 1, Figs. 2, 3)
Neogloboquadrina inglei Kucera and Kennett, 2000, (Plate 1, Figs. 1–13) [1].
Test relatively large, almost exclusively dextrally coiled, rounded, lobulate in both axial and equatorial peripheries. Four to three and a half globular chambers in the final whorl, slowly expanding in size giving the test somewhat quadrate appearance. Aperture narrow umbilical–extraumbilical bordered with thin lip. Populations of this form at Site 1150 frequently exhibit a distinct rim-like lip.
Neogloboquadrina incompta (Cifelli) (Plate 2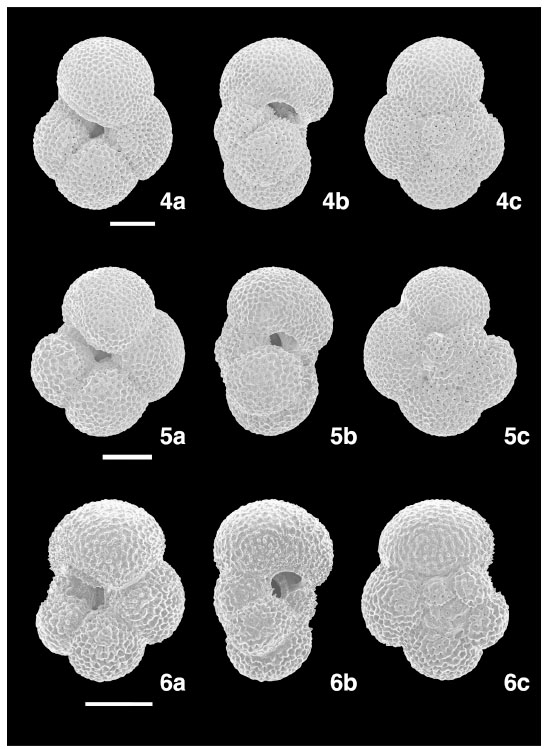 , Figs. 4–6)
, Figs. 4–6)

Globigerina incompta Cifelli, 1961, (Plate 4, Figs. 1–7) [9].
Test relatively small, dominantly dextrally coiled, lobulate equatorial periphery, gently compressed axial periphery. Four to four and a half globular chambers in the final whorl, moderately increasing in size as added, except for the last 3–4 chambers which are added rapidly. Aperture umbilical–extraumbilical, a rather low arch with a thin lip. Scanning electron micrographs of this species show the typical neogloboquadrinid wall structure without spines.
Neogloboquadrina asanoi (Maiya, Saito and Sato) (Plate 3, Figs. 7–9)

Globoquadrina asanoi Maiya, Saito and Sato, 1976, (Plate 3, Figs. 1–3) [10].
Test large, almost exclusively dextrally coiled, moderately lobulate equatorial periphery, broadly rounded axial periphery. Four globular chambers in the final whorl, gradually increasing in size as added, protruding conical arrangement of early whorls. Aperture interiomarginal, umbilical, a low arch with no apparent modifications.
ACKNOWLEDGEMENTS
We express our gratitude to Dr. M. Kucera, Dr. S. Maiya, and the late Dr. R. Cifelli for kindly providing the topotypes of Neogloboquadrina inglei, Globoquadrina asanoi, and Globigerina incompta, respectively. We thank Ms. C. Morishita for assistance with sample treatments. This research was supported in part by a Grant-in-Aid for Scientific Research (A) from JSPS, No. 17204043 to M. Oda, and by the 21st Century Center-Of-Excellence Program “Advanced Science and Technology Center for the Dynamic Earth” of Tohoku University.