- Home
- About Journals
-
Information for Authors/ReviewersEditorial Policies
Publication Fee
Publication Cycle - Process Flowchart
Online Manuscript Submission and Tracking System
Publishing Ethics and Rectitude
Authorship
Author Benefits
Reviewer Guidelines
Guest Editor Guidelines
Peer Review Workflow
Quick Track Option
Copyediting Services
Bentham Open Membership
Bentham Open Advisory Board
Archiving Policies
Fabricating and Stating False Information
Post Publication Discussions and Corrections
Editorial Management
Advertise With Us
Funding Agencies
Rate List
Kudos
General FAQs
Special Fee Waivers and Discounts
- Contact
- Help
- About Us
- Search




The Open Virology Journal
(Discontinued)
ISSN: 1874-3579 ― Volume 15, 2021
Exploration of New Sites in Adenovirus Hexon for Foreign Peptides Insertion
Satyender Hansra, Sujit Pujhari , Alexander N Zakhartchouk*
Abstract
Adenoviral vectors are now being explored as vaccine carriers to prevent infectious diseases in humans and animals. There are two strategies aimed at the expression of a vaccine antigen by adenoviral vectors. The first includes an insertion of the foreign gene expression cassette into the E1 region. The second strategy is antigen incorporation into the viral capsid proteins. To extend this methodology, we have searched for new sites at the human adenovirus serotype 5 major capsid protein hexon for a vaccine antigen insertion. To this end, we utilized sites in the hexon hypervariable region (HVR) 7, 8 and 9 to display a 15-mer peptide containing the main neutralizing epitope of porcine reproductive and respiratory syndrome virus. However, we could not rescue the viruses with the insertions of the peptide into HVR 8 and 9, consistent with the viruses being unable to tolerate insertions at these sites. In contrast, the virus with the insertion of the peptide in HVR 7 was viable - growing well in cell culture and the inserted peptide was exposed on the virion surface.
Article Information
Identifiers and Pagination:
Year: 2015Volume: 9
First Page: 1
Last Page: 6
Publisher Id: TOVJ-9-1
DOI: 10.2174/1874357901509010001
Article History:
Received Date: 7/4/2015Revision Received Date: 28/4/2015
Acceptance Date: 30/4/2015
Electronic publication date: 28/5/2015
Collection year: 2015
open-access license: This is an open access article licensed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted, non-commercial use, distribution and reproduction in any medium, provided the work is properly cited.
* Address correspondence to this author at the Vaccine and Infectious Disease Organization-International Vaccine Center (VIDO-InterVac), University of Saskatchewan, 120 Veterinary Road, Saskatoon, SK S7N 5E3, Canada; Tel: 1-306-966-1570; Fax: 1-306-966-7478; E-mail: alex.zak@usask.ca
| Open Peer Review Details | |||
|---|---|---|---|
| Manuscript submitted on 7-4-2015 |
Original Manuscript | Exploration of New Sites in Adenovirus Hexon for Foreign Peptides Insertion | |
BACKGROUND
Human adenovirus type 5 (hAd5) vector has been shown to be an excellent vaccine delivery system in humans and animals [1Lasaro MO, Ertl HC. New insights on adenovirus as vaccine vectors Mol Ther 2009; 17: 1333-9., 2Ferreira TB, Alves PM, Aunins JG, Carrondo MJ. Use of adenoviral vectors as veterinary vaccines Gene Ther 2005; 12: S73-83.]. The replication-defective (E1-deleted) and replication-competent (E3-deleted) hAd5 have both been used as vaccine delivery vectors. For biosafety concerns, a replication-defective hAd5 is considered to be more suitable for vaccination. Replication-defective vectors may also have deletion in the E3 region to increase the amount of foreign DNA that can be inserted. Conventional strategy for the expression of a vaccine antigen by adenoviral vector includes insertion of the foreign gene expression cassette into the E1 region. This antigen is expressed as transgene after the infection of permissive cells with recombinant adenovirus.
Apparent drawbacks of the conventional transgene expression of antigen include an inability of the hAd5-based vectors to produce a potent humoral response against certain antigens and, in some cases, the inability of the vector to express a foreign gene [3Kheyar A, Jabrane A, Zhu C , et al. Alternative codon usage of PRRS virus ORF5 gene increases eucaryotic expression of GP(5):glycoprotein and improves immune response in challenged pigs Vaccine 2005; 23: 4016-22.]. To overcome such hurdles, the “antigen capsid-incorporation” strategy was developed [4Matthews QL. Capsid-incorporation of antigens into adenovirus capsid proteins for a vaccine approach Mol Pharm 2011; 8: 3-11.]. Incorporating an immunogenic peptide into the hAd5 capsid offers several potential advantages. Importantly, the processing of capsid-incorporated antigen via the exogenous pathway could result in a strong humoral response, similar to that generated by native hAd5 capsid proteins, and this strategy may also be useful in boosting antigen-specific antibody immune responses.
Adenoviral capsid consists of the hexon, penton base and fiber [5Rux JJ, Burnett RM. Adenovirus structure Hum Gene Ther 2004; 15: 1167-76.]. Antigenic epitopes could be incorporated into these proteins as well as into the minor capsid protein IX [6Coughlan L, Alba R, Parker AL , et al. Tropism-modification strategies for targeted gene delivery using adenoviral vectors Viruses 2010; 2: 2290-355.]. Importantly, hexon is the most abundant of the capsid proteins, accounting for more than 83% of the capsid protein [7Rux JJ, Kuser PR, Burnett RM. Structural and phylogenetic analysis of adenovirus hexons by use of high-resolution x-ray crystallographic, molecular modeling, and sequence-based methods J Virol 2003; 77: 9553-66.]. Furthermore, hexon is shown to be a vaccine adjuvant [8Molinier-Frenkel V, Lengagne R, Gaden F , et al. Adenovirus hexon protein is a potent adjuvant for activation of a cellular immune response J Virol 2002; 76: 127-35.].
Early analysis of the hexon protein sequences revealed that, in addition to the conserved regions, there were 7 discrete hypervariable regions (HVRs) [9Crawford-Miksza L, Schnurr DP. Analysis of 15 adenovirus hexon proteins reveals the location and structure of seven hypervariable regions containing serotype-specific residues J Virol 1996; 70: 1836-44.]. In a subsequent study, they are reclassified as regions 1 to 9 [7Rux JJ, Kuser PR, Burnett RM. Structural and phylogenetic analysis of adenovirus hexons by use of high-resolution x-ray crystallographic, molecular modeling, and sequence-based methods J Virol 2003; 77: 9553-66.]. These HVRs do not appear to be involved in the binding of any other viral proteins, and the loops at the top of the HVRs are the most amenable to modifications. It was also shown that short heterologous peptides can be incorporated within the HVRs of hexon without affecting the viability of the virus [10Wu H, Han T, Belousova N , et al. Identification of sites in adenovirus hexon for foreign peptide incorporation J Virol 2005; 79: 3382-90., 11Matthews QL, Yang P, Wu Q , et al. Optimization of capsid-incorporated antigens for a novel adenovirus vaccine approach Virol J 2008; 5: 98.]. Incorporating antigens into hexon is applicable towards vaccination for several pathogens including the poliovirus, pseudomonas, B. anthracis, influenza, malaria and HIV [12Worgall S, Krause A, Qiu J, Joh J, Hackett NR, Crystal RG. Protective immunity to pseudomonas aeruginosa induced with a capsid-modified adenovirus expressing, P aeruginosa OprF J Virol 2007; 81: 13801-8.-17McConnell MJ, Danthinne X, Imperiale MJ. Characterization of a permissive epitope insertion site in adenovirus hexon J Virol 2006; 80: 5361-70.]. In selected studies, protective immune responses in a mouse model are documented [12Worgall S, Krause A, Qiu J, Joh J, Hackett NR, Crystal RG. Protective immunity to pseudomonas aeruginosa induced with a capsid-modified adenovirus expressing, P aeruginosa OprF J Virol 2007; 81: 13801-8., 16Shiratsuchi T, Rai U, Krause A, Worgall S, Tsuji M. Replacing adenoviral vector HVR1 with a malaria B cell epitope improves immunogenicity and circumvents preexisting immunity to adenovirus in mice J Clin Invest 2010; 120: 3688-701.].
In hAd5, HVR 1 through 6, and 8 have been used for the incorporation of foreign peptides [10Wu H, Han T, Belousova N , et al. Identification of sites in adenovirus hexon for foreign peptide incorporation J Virol 2005; 79: 3382-90., 18Gu L, Li ZC, Krendelchtchikov A, Krendelchtchikova V, Wu H, Matthews QL. Using multivalent adenoviral vectors for HIV vaccination PLoS One 2013; 8: e60347.]. In this study, we explored different sites in HVRs 7, 8 and 9 of hAd5 hexon for the insertion of a peptide corresponding to the porcine reproductive and respiratory syndrome virus (PRRSV) main neutralizing epitope B [19Ostrowski M, Galeota JA, Jar AM, Platt KB, Osorio FA, Lopez OJ. Identification of neutralizing and nonneutralizing epitopes in the porcine reproductive and respiratory syndrome virus GP5 ectodomain J Virol 2002; 76: 4241-50.] of the GP5 protein. Our results provide potentially applicable information for adenovirus-vectored vaccine development involving such hexon modifications, particularly in HVR 7.
RESULTS AND DISCUSSION
Construction of Hexon-Modified Viruses
In order to access new sites in hAd5 hexon, tailored for the incorporation of foreign peptide sequences, we genetically inserted a 15 amino acid (aa) residue peptide into HVR7, HVR8 or HVR9 region of the hAd5 hexon. The peptide consisted of embedding a 9 aa residue sequence containing the PRRSV major neutralizing epitope flanked by the LGS spacer sequences (Fig. 1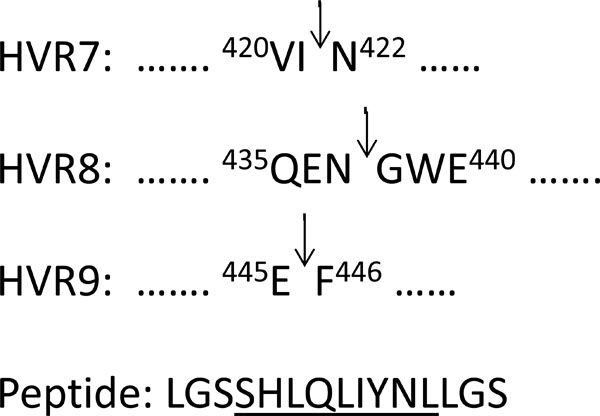 ).
).

Insertion of the DNA encoding peptide into the hAd5 hexon gene was achieved by PCR followed by bacterium-based homologous recombination. The linearized DNA of the plasmid containing the right portion of the hAd5 genome with modified hexon and the 1878 bp deletion in the E3 region was co-transfected into human embryo kidney (HEK) 293 cells together with a plasmid containing the left portion of the hAd5 genome with an E1-deletion. Homologous recombination in HEK 293 cells led to generation of the recombinant virus, termed Ad5HVR7epB. Multiple attempts to rescue the other two recombinant viruses (Ad5HVR8epB and Ad5HVR9epB) were not successful. This inability to rescue recombinant viruses with the foreign peptide insertion into HVR8 and HVR9 of hexon suggested that the insertion interferes with the formation of viral particles.
Interestingly, in a previous study [10Wu H, Han T, Belousova N , et al. Identification of sites in adenovirus hexon for foreign peptide incorporation J Virol 2005; 79: 3382-90.], hexon-modified virus with incorporation of the His6 peptide into HVR8 was reported to be viable. Our result with HVR8 is contradictory, evidently due to the differences in the peptides sequences and their lengths. Further, while the two studies are made with changes within HVR8, the specific locations of the incorporations differ as well. Notably our study uses a foreign sequence inserted between Asn437 and Gly438 (Fig. 1), while Wu et al. [10Wu H, Han T, Belousova N , et al. Identification of sites in adenovirus hexon for foreign peptide incorporation J Virol 2005; 79: 3382-90.] placed His6 between Pro431 and Trp439 substituting for 7 aa residues of hexon.
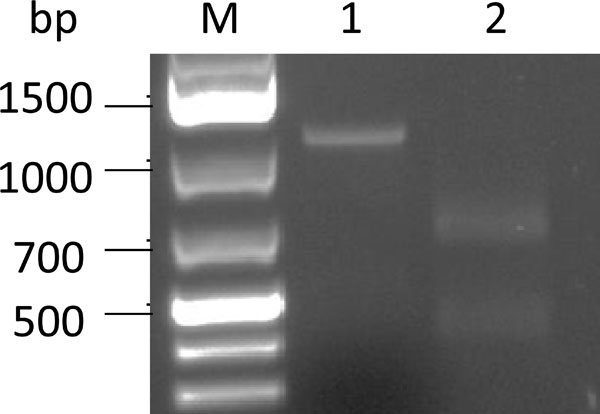
To corroborate the insertion of the peptide encoding DNA into the hexon gene of hAd5, the viral genome fragment was amplified using PCR from the viral DNA extracted from Ad5HVR7epB-infected cells, and the PCR product was digested with AvrII since the peptide encoding DNA contained the single AvrII site. Predictably, a 1053 bp PCR fragment was amplified from Ad5HVR7epB DNA (Fig. 2 , lane 1), and its site specific cleavage by AvrII provided two fragments: 650 bp and 403 bp (Fig. 2, lane 2). The results of this analysis suggested that recombinant Ad5HVR7epB contained the epitope B encoding sequence in the HVR7 of hexon.
, lane 1), and its site specific cleavage by AvrII provided two fragments: 650 bp and 403 bp (Fig. 2, lane 2). The results of this analysis suggested that recombinant Ad5HVR7epB contained the epitope B encoding sequence in the HVR7 of hexon.
Growth of the Hexon-Modified Virus in Cell Culture
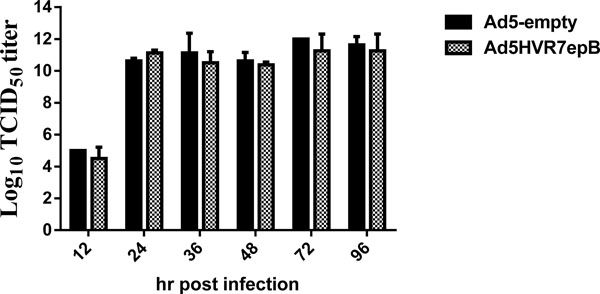
The growth kinetics of the recombinant virus Ad5HVR7epB was analyzed in HEK 293 cells. Cultures of the cells were infected with Ad5HVR7epB or Ad5-empty (E1-deleted and partially E3-deleted hAd5 with unmodified hexon), and the cells were harvested at 12, 24, 36, 48, 72 and 96 hr post infection intervals. Virus in each sample was released by freeze-thawing and titered on monolayers of HEK 293 cells. Fig. (3 ) shows the insertion of the peptide in HVR7, resulting in similar virus yields to unmodified virus with maximal titers ranging from 2 X 1011 to 1012 TCID50 per ml in a culture of 2 X 106 cells.
) shows the insertion of the peptide in HVR7, resulting in similar virus yields to unmodified virus with maximal titers ranging from 2 X 1011 to 1012 TCID50 per ml in a culture of 2 X 106 cells.
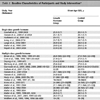
We also determined the viral particle/infectious particle (VP/IP) ratio for CsCl gradient purified preparations of the control virus Ad5-empty, as well as the hexon-modified virus Ad5HVR7epB. A 2.7 fold increase was observed in the VP/IP ratio of Ad5HVR7epB preparations as compared to the unmodified control Ad5-empty (Table 2). A similar observation is reported for hexon-modified hAd5 with peptide incorporation into HVR2 or 5 [11Matthews QL, Yang P, Wu Q , et al. Optimization of capsid-incorporated antigens for a novel adenovirus vaccine approach Virol J 2008; 5: 98.]. A typical VP/IP ratio of unmodified adenoviruses ranges from 10 to 44 [20Both GW, Cameron F, Collins A, Lockett LJ, Shaw J. Production and release testing of ovine atadenovirus vectors Methods Mol Med 2007; 130: 69-90.].
Presentation of the Foreign Peptide on Virion Surface
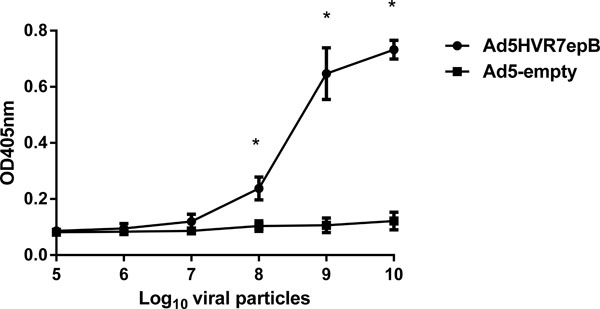
We next examined whether PRRSV epitope was presented on the surface of the virion, using an ELISA binding assay. Different amounts of CsCl gradient purified viruses (Ad5HVR7epB and Ad5-empty) were immobilized on the wells of 96-well plates and incubated with peptide antisera. The binding of antibodies to the virions was detected with alkaline phosphatase (AP)-conjugated secondary antibody, followed by AP color reaction development. Evidently, the anti-peptide antibodies bind well to AdHVR7epB virions and not to Ad5-empty (Fig. 4 ). This suggests that the PRRSV epitope inserted into HVR7 of hexon is exposed on the virion surface.
). This suggests that the PRRSV epitope inserted into HVR7 of hexon is exposed on the virion surface.
While a previous study showed that epitopes were exposed on the virion surface when incorporated in HVR1, HVR2 and HVR5, and not exposed in HVR3 and HVR6, as well as poorly exposed when a peptide was placed in HVR8 [10Wu H, Han T, Belousova N , et al. Identification of sites in adenovirus hexon for foreign peptide incorporation J Virol 2005; 79: 3382-90.]. In the current study, we find a new site in HVR7 for incorporation of foreign peptide into hAd5 hexon, and determine that the peptide was also exposed on the virion surface making it readily accessible for antibody binding and be potentially useful for vaccination.
CONCLUSION
Herein, we report a novel adenovirus vector (Ad5HVR7epB) with the insertion of the PRRSV main neutralizing epitope B in 3 different HVR regions of the major capsid protein hexon. While the insertion of the peptide into HVRs 8 and 9 were not tolerated by the adenovirus, the 9-mer epitope placed within HVR7 is determined to be exposed on the virion surface.
METHODS
Construction of the Plasmids pUC-Ad5hex-HVR7epB, pUC-Ad5hex-HVR8epB and pUC-Ad5hex-HVR9epB
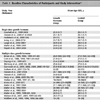
Plasmid pUC-Ad5-hex was used as a template for PCR and as a cloning vector. This plasmid contained a 5.2 kb DNA fragment of hAd5 genome encoding a portion of the hexon gene. To insert the DNA sequence encoding the main neutralizing epitope B of PRRSV in HVR7 and obtain the DNA fragment for cloning, four consequent PCRs were performed using Phusion high-fidelity DNA Polymerase (NEB). First, a 1 kb fragment was amplified using primers BL7F1 and Nhe-R, and pUC-Ad5-hex as a template. Second, a 1 kb fragment was amplified using primers BLF2 and Nhe-R, and the product of PCR 1 as a template. Third, a 690 bp fragment was amplified using primers BL7R and Nde-F, and pUC-Ad5-hex as a template. Fourth, a 1.7 kb fragment was amplified using primers Nde-F and Nhe-R, and the mixture of the products of PCR 2 and 3 from above as a template. The final 1.7 kb PCR product was digested with NdeI and NheI and cloned into NdeI and NheI sites of pUC-Ad5-hex, creating pUC-Ad5hex-HVR7epB. Similar strategy was used to construct pUC-Ad5hex-HVR8epB and pUC-Ad5hex-HVR9epB. Specific sequences of the primers are presented in Table 1.
Construction of the Plasmids pH5R-hexHVR7epB, pH5R-hexHVR8epB and pH5R-hexHVR9epB
The plasmid pH5R-hexHVR7epB was generated by homologous DNA recombination in E. coli BJ5183 [21Reddy PS, Ganesh S, Hawkins L, Idamakanti N. Generation of recombinant adenovirus using the Escherichia coli BJ5183 recombination system Methods Mol Med 2007; 130: 61-8.] between a 2.6 kb DNA fragment excised by KpnI and NdeI from pUC-Ad5hex-HVR7epB and BamHI-linearized pH5R [22Zakhartchouk AN, Viswanathan S, Mahony JB, Gauldie J, Babiuk LA. Severe acute respiratory syndrome coronavirus nucleocapsid protein expressed by an adenovirus vector is phosphorylated and immunogenic in mice J Gen Virol 2005; 86: 211-5.]. Similar strategy was used to construct pH5R-hexHVR8epB and pH5R-hexHVR9epB. Fig. (1) shows the sites of the insertion of the peptide in the hAd5 hexon protein sequence.
Rescue of Recombinant Adenoviruses
Human embryonic kidney (HEK) 293 cells were transfected with PacI-digested DNAs of the plasmids pH5R-hexHVR7epB, pH5R-hexHVR8epB or pH5R-hexHVR9epB and PacI-digested pH5-L [22Zakhartchouk AN, Viswanathan S, Mahony JB, Gauldie J, Babiuk LA. Severe acute respiratory syndrome coronavirus nucleocapsid protein expressed by an adenovirus vector is phosphorylated and immunogenic in mice J Gen Virol 2005; 86: 211-5.] using Lipofectamine-2000 (Invitrogen). Homologous recombination in the transfected cells led to generation of the recombinant adenovirus, and the virus was detected by developing cytopathic effect (CPE) in the cell monolayer. If CPE was not detected by the 14th day after transfection, the cells and the cell culture media were collected and freeze-thawed. This material was used to infect a fresh monolayer of HEK 293 cells.
The virus was amplified by passaging in HEK 293 cells and purified by CsCl gradient centrifugation [23Tollefson AE, Kuppuswamy M, Shashkova EV, Doronin K, Wold WS. Preparation and titration of CsCl-banded adenovirus stocks Methods Mol Med 2007; 130: 223-35.]. The concentration of virus particles (VP) in purified virus preparations was determined by measuring the optical density at 260 nm as previously described [24Mittereder N, March KL, Trapnell BC. Evaluation of the concentration and bioactivity of adenovirus vectors for gene therapy J Virol 1996; 70: 7498-509.], whereas numbers of infectious particles (IP) were determined by the tissue culture infective dose 50 (TCID50) method [25Zakhartchouk AN, Wu Q, Tikoo SK. Construction of capsid-modified recombinant bovine adenovirus type 3 Methods Mol Med 2007; 130: 91-106.].
Analysis of Virus Growth
HEK 293 cells grown in wells of 6-well plates were infected with Ad5-empty or Ad5HVR7epB viruses at a multiplicity of infection (m.o.i.) of 5. Infected cells were harvested at the indicated time intervals post-infection. Cells were lysed in the growth medium by freezing-thawing three times. The titres of infectious viral progeny were determined by TCID50 method in monolayers of HEK 293 cells.
Isolation of Adenoviral DNA from Infected Cell Lysates and PCR
HEK 293 cells grown in T75 flasks were infected with the virus Ad5HVR7epB. After 48 hours post infection, when full CPE was observed, the cells and cell culture media were harvested followed by 3 cycles of freezing and thawing. Cell debris was removed by centrifugation at 1,500 rpm for 5 min, and the supernatant (275 μl) was used for viral DNA isolation. Briefly, the supernatant was incubated with 10 μl of DNase I (10 mg/ml) at 370C for 30 min, followed by addition of 6 μl of 0.5 M ethylenediaminetetraacetic acid (EDTA) (pH 8.0), 7.5 μl of 20 % sodium dodecyl sulfate (SDS), 1.5 μl of Proteinase K (20 mg/ml) and incubation at 500C for 1 h. Total DNA was further extracted using a GeneClean Spin Kit (MP Biomedicals) according to the manufacturer’s instructions. The isolated DNA was subjected to PCR using Fermentas (2X) PCR Master Mix and primers (5’-ATCATGCAGCTGGGAGAGTC-3’ and 5’-CATTGCCCAGCAACATTGAG-3’) that were designed to anneal in the sites flanking the hexon HVRs. Amplification product was digested by AvrII and size-separated by electrophoresis in 1 % agarose gel.
Production of Peptide-Specific Polyclonal Antibodies
The peptide, containing amino acid residues of the PRRSV neutralizing epitope B (SHLQLIYNL), was synthesized by GenScript Inc. The peptide was conjugated to keyhole limpet hemocyanin (KLH) or bovine serum albumin (BSA) as carrier molecules. Two New Zealand White rabbits were immunized with the conjugated peptide (500 μg/rabbit) emulsified with Freund’s Complete Adjuvant (FCA) followed by two injections of conjugated peptide (300 μg/rabbit) in Freund’s incomplete adjuvant (FIA) at four weeks apart. Serum was collected ten days after the third injection and tested for ability to bind to the peptide by ELISA.
ELISA
The enzyme-linked immunosorbent assay (ELISA) binding assay was used to test if the peptide incorporated into the hexon protein is accessible to anti-peptide antibody at the virion level. Different amounts of purified virions ranging from 105 to 1010 viral particles were immobilized on the wells of a 96-well plate (IMMULON, Thermo Scientific) by overnight incubation in 100 ul/well of 100 mM carbonate buffer (pH 9.5) at 4 0C. After washing with PBS, containing 0.05% Tween 20, the immobilized virus was incubated with 1:500 diluted anti-peptide rabbit serum for 4 h at 25 0C, followed by AP-conjugated goat anti-rabbit antibody (Cedarlane) incubation. Color reaction was performed with p-nitrophenyl phosphate (Sigma-Aldrich), and optical density at 405 nm was measured with a microplate reader.
ABBREVIATIONS
CONFLICT OF INTEREST
The authors confirm that this article content has no conflict of interest.
ACKNOWLEDGEMENTS
This work was supported by grants from the Canadian Swine Health Board and the Saskatchewan Ministry of Agriculture. We thank the staff of the VIDO-InterVac Animal Care Unit for immunizations of rabbits. This paper was published with the permission of the Director of VIDO-InterVac, journal series no. 719.