- Home
- About Journals
-
Information for Authors/ReviewersEditorial Policies
Publication Fee
Publication Cycle - Process Flowchart
Online Manuscript Submission and Tracking System
Publishing Ethics and Rectitude
Authorship
Author Benefits
Reviewer Guidelines
Guest Editor Guidelines
Peer Review Workflow
Quick Track Option
Copyediting Services
Bentham Open Membership
Bentham Open Advisory Board
Archiving Policies
Fabricating and Stating False Information
Post Publication Discussions and Corrections
Editorial Management
Advertise With Us
Funding Agencies
Rate List
Kudos
General FAQs
Special Fee Waivers and Discounts
- Contact
- Help
- About Us
- Search




The Open Bioactive Compounds Journal
(Discontinued)
ISSN: 1874-8473 ― Volume 9, 2020
Allantoin as A Biomarker of Inflammation in an Inflammatory Bowel Disease Mouse Model: NMR Analysis of Urine
Philippa A. Dryland1, ¥, Donald R. Love2, ¥, Michael F. Walker3, Yvonne Dommels4, ¥, #, Christine Butts4, ¥, Daryl Rowan5, ¥, Nicole C. Roy6, ¥, Nuala Helsby7, Brian L. Browning1, ¥, Shuotun Zhu8, ¥, Brent R. Copp3 , Lynnette R. Ferguson1, ¥, *
Abstract
Crohn’s disease (CD) is an Inflammatory Bowel Disease (IBD) that is characterised by destructive inflammation of the intestinal wall. Current methods for determining inflammation of the bowel are costly, time consuming and can cause discomfort to the patients. In order to address these problems, biomarker analysis of more accessible tissues is receiving increasing attention. Oxidative stress has been implicated in the promotion of inflammation. Allantoin has recently been reported as a biomarker for oxidative stress in human serum and urine. This paper investigates allantoin as a biomarker of inflammation in a mouse model of CD. Proton nuclear magnetic resonance (1H NMR) spectroscopy was used to analyse allantoin in urine from the mdr1a -/- mouse which is a model of CD. The data show that the levels of allantoin are strongly correlated with histological injury scores of mouse colonic tissue samples. Allantoin appears to be a useful biomarker of gut inflammation, involving oxidative stress, in a mouse model of CD and may be a potential biomarker in human CD studies
Article Information
Identifiers and Pagination:
Year: 2008Volume: 1
First Page: 1
Last Page: 6
Publisher Id: TOBCJ-1-1
DOI: 10.2174/1874847300801010001
Article History:
Received Date: 15/1/2008Revision Received Date: 4/4/2008
Acceptance Date: 1/5/2008
Electronic publication date: 3/6/2008
Collection year: 2008
open-access license: This is an open access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.5/), which permits unrestrictive use, distribution, and reproduction in any medium, provided the original work is properly cited.
* Address correspondence to this author at the Department of Nutrition, University of Auckland, Private Bag 92019, Auckland Mail Centre, Auckland 1142, New Zealand; Tel: +649 373 7599, ext 86372; E-mail: l.ferguson@auckland.ac.nz
| Open Peer Review Details | |||
|---|---|---|---|
| Manuscript submitted on 15-1-2008 |
Original Manuscript | Allantoin as A Biomarker of Inflammation in an Inflammatory Bowel Disease Mouse Model: NMR Analysis of Urine | |
INTRODUCTION
Crohn’s disease (CD) is an Inflammatory Bowel Disease (IBD) that is characterised by destructive inflammation of the gastrointestinal tract. The disease is believed to be caused by a disproportionate immune response to environmental factors in people with a genetic predisposition [1Gonzalez C, Almaraz L, Obeso A, Rigual R. Physiol Rev 1994; 74: 829.
[PMID: 7938227] ,2Gonzalez C, Lopez-Lopez JR, Obeso A, Perez-Garcia MT, Rocher A. Respir Physiol 1995; 102: 137.
[http://dx.doi.org/10.1016/0034-5687(95)00069-0] ]. This immune response has a key role in the initiation and maintenance of inflammation which characterises CD patho-genesis [3Riesco-Fagundo AM, Perez-Garcia MT, Gonzalez C, Lopez-Lopez JR. Circ Res 2001; 89: 430.
[http://dx.doi.org/10.1161/hh1701.095632] [PMID: 11532904] ]. The symptoms of CD are diverse depending on the location of inflammation, but general symptoms include abdominal pain, diarrhoea and rectal bleeding [4Lahiri S, Roy A, Baby SM, Hoshi T, Semenza GL, Prabhakar NR. Prog Biophys Mol Biol 2006; 91: 249.
[http://dx.doi.org/10.1016/j.pbiomolbio.2005.07.001] [PMID: 16137743] ]. At present, several clinical investigations are required to make an accurate diagnosis of CD. These investigations include invasive endoscopy, serological testing and radiological testing. They are time consuming, costly and can cause discomfort to the patients [5Semenza GL. Hydroxylation of HIF-1: oxygen sensing at the molecular level Physiology (Bethesda) 2004; 19: 176.
[http://dx.doi.org/10.1152/physiol.00001.2004] [PMID: 15304631] ]. As a complement to these diagnostic methods, biomarkers offer the prospect of providing an assessment of underlying biological events that occur as a consequence of disease progression [6Semenza GL. J Appl Physiol 2004; 96: 1170.], and importantly do not require invasive procedures.
In the context of IBD oxidative stress could offer an appropriate entry point in identifying useful biomarkers.
Oxidative stress has been implicated in the promotion of inflammation due to the formation of peroxynitrite and downstream activation of transcriptional factors such as NF-κB [7Baby SM, Roy A, Mokashi AM, Lahiri S. Histochem Cell Biol 2003; 120: 343.
[http://dx.doi.org/10.1007/s00418-003-0588-2] [PMID: 14600837] ,8Roy A, Volgin DV, Baby SM, Mokashi A, Kubin L, Lahiri S. Neurosci Lett 2004; 363: 229.
[http://dx.doi.org/10.1016/j.neulet.2004.03.073] [PMID: 15182949] ]. Critically, allantoin has been reported as a sensitive marker of oxidative stress [9Tipoe GL, Fung ML. Respir Physiol Neurobiol 2003; 138: 143.
[http://dx.doi.org/10.1016/S1569-9048(03)00188-5] ,10Milkiewicz M, Pugh CW, Egginton S. J Physiol 2004; 560: 21.
[http://dx.doi.org/10.1113/jphysiol.2004.069757] [PMID: 15319416] [PMCID: PMC1665195] ]. In most mammals uric acid is converted to allantoin by urate oxidase; however, in humans the urate oxidase gene is not expressed due to several “nonsense” mutations and uric acid is converted to allantoin via reactive oxygen species as shown in Fig. (1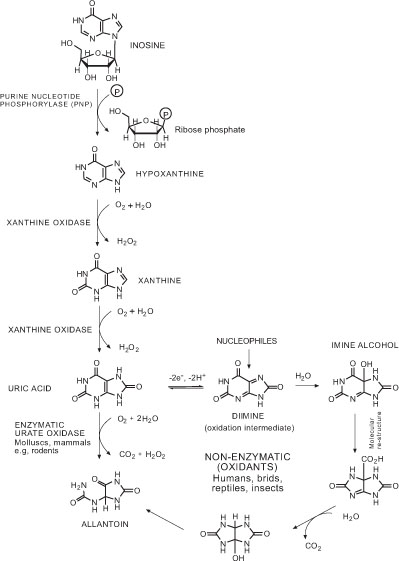 ) [9Tipoe GL, Fung ML. Respir Physiol Neurobiol 2003; 138: 143.
) [9Tipoe GL, Fung ML. Respir Physiol Neurobiol 2003; 138: 143.
[http://dx.doi.org/10.1016/S1569-9048(03)00188-5] ,11Roy A, Li J, Baby SM, Mokashi A, Buerk DG, Lahiri S. Respir Physiol Neurobiol 2004; 141: 115.
[http://dx.doi.org/10.1016/j.resp.2004.03.010] [PMID: 15239962] ]. Taken together, allantoin could be a likely candidate biomarker of CD.

Mouse models of CD offer a tractable system to identify appropriate biomarkers for further validation in humans. Of the mouse models that have been reported in the literature, mice with targeted deletions of the mdr1a gene spontaneously develop intestinal inflammation in specific pathogen-free conditions [12Vicario I, Rigual R, Obeso A, Gonzalez C. Am J Physiol Cell Physiol 2000; 278: C490.
[PMID: 10712237] ]. A recent study by Dommels et al. (2007) reported spontaneous development of inflammation between 16 and 25 weeks of age in the mdr1a-/- mice. The lesions observed in these mice were transmural and discontinuous, located in the colon and small intestine, similar to those in human IBD, specifically CD [13Nurse CA. Auton Neurosci 2005; 120: 1.
[http://dx.doi.org/10.1016/j.autneu.2005.04.008] [PMID: 15955746] ,14Lahiri S, Baby SM, Roy A. Curr Respir Med Rev 2006; 2: 393.
[http://dx.doi.org/10.2174/157339806778777221] ].
On the basis of the above, an NMR spectroscopy approach was used to [15Prasad M, Fearon IM, Zhang M, Laing M, Vollmer C, Nurse CA. J Physiol 2001; 537: 667.
[http://dx.doi.org/10.1113/jphysiol.2001.012836] -18Iturriaga R, Villanueva S, Mosqueira M. J Appl Physiol 2000; 89: 1005.] study metabolites in the urine of mdrla gene targeted mutant mice. The objective of this study was to investigate allantoin as a viable biomarker of inflammation in an IBD mouse model.
MATERIALS AND METHODS
Animals
In this study urine from 46 male mice was tested using 1H NMR analyses. The urine was collected from a mouse study recently performed by Dommels et al. (2007) [13Nurse CA. Auton Neurosci 2005; 120: 1.
[http://dx.doi.org/10.1016/j.autneu.2005.04.008] [PMID: 15955746] ]. Twenty three mice were a control strain (FVB), and the remaining 23 were homozygous for the targeted mutation of the mdr1a gene. All of the mice were sourced from Taconic Farms Inc. (Germantown, NY, USA) at four to five weeks of age. Mice were individually housed in shoe-box style cages with free access to water and a plastic tube for environmental enrichment. The mice were fed an AIN-76A powdered diet. The intestinal samples were obtained from mice between 12 to 29 weeks of age which corresponds to the period of intestinal inflammation in mdr1a gene targeted mutant mice. The intestine was removed swiftly, a lengthwise incision was made to open and the intestine was flushed with a 0.9% NaCl solution to remove traces of digesta. The intestine was subdivided into 4 sections, duodenum, jejunum, ileum and colon. Subsections of these were placed into formalin (10% neutral buffered) and stored at room temperature for histological analysis [13Nurse CA. Auton Neurosci 2005; 120: 1.
[http://dx.doi.org/10.1016/j.autneu.2005.04.008] [PMID: 15955746] ,19Prabhakar NR. Respir Physiol 1999; 115: 161.
[http://dx.doi.org/10.1016/S0034-5687(99)00019-5] ]. The urine samples were collected from mice housed in metabolic cages five days prior to the harvesting of tissues as described by Dommels et al. (2007) [13Nurse CA. Auton Neurosci 2005; 120: 1.
[http://dx.doi.org/10.1016/j.autneu.2005.04.008] [PMID: 15955746] ]. The experimental procedures for this trial were reviewed and approved by the Crown Research Institute Animal Ethics Committee in Palmerston North, New Zealand according to the New Zealand Animal Welfare Act (1999).
HISTOLOGICAL ANALYSIS
All histological analysis was performed blinded to the strain and treatment of the mouse. Two intact pieces from different areas of each intestinal section (duodenum, jejunum, ileum or colon) were fixed in 10% (v/v) neutral buffered formalin and embedded in a paraffin block. Five micron sections were stained with haematoxylin and eosin for light microscopic examination in order to score three principal aspects of inflammation [20Wang ZZ, Dinger BG, Stensaas LJ, Fidone SJ. Biol Signals 1995; 4: 109.
[http://dx.doi.org/10.1159/000109430] [PMID: 8750936] , 21Wang ZZ, Stensaas LJ, Dinger BG, Fidone SJ. Neuroscience 1995; 65: 217.
[http://dx.doi.org/10.1016/0306-4522(94)00437-A] ]. These aspects included inflammatory lesions (mononuclear cell infiltration, neutrophil infiltration, eosinophil infiltration, plasmocyte infiltration, fibrin exudation and lymphangiectasis), tissue destruction (enterocyte loss, ballooning degeneration, edema and mucosal atrophy) and tissue reparation (hyperplasia, angiogenesis, granulomas and fibrosis). A rating score between 0 (no change from normal tissue) and 3 (lesions that involved most areas and all layers of the intestinal section including mucosa, muscle and omental fat) was given for each principal aspect of inflammation. A total histological injury score (HIS) was calculated for each intestinal section. The HIS comprised the sum of the inflammatory lesions score (multiplied by two to give weight to this predominant biological aspect of inflammation), and the tissue destruction and tissue reparation scores [13Nurse CA. Auton Neurosci 2005; 120: 1.
[http://dx.doi.org/10.1016/j.autneu.2005.04.008] [PMID: 15955746] ].
NMR ANALYSIS
Two sets of NMR data were used for analysis. The first set of urine samples (13 FVB mice and 13 homozygous mdr1a gene targeted mutant mice) were aliquoted into 500 µl samples that were added to 5 mm NMR tubes. The second set of urine samples (10 FVB mice and 10 homozygous mdr1a gene targeted mutant mice) were prepared using a protocol described by Stanley et al. [18Iturriaga R, Villanueva S, Mosqueira M. J Appl Physiol 2000; 89: 1005.]. This protocol involved adding a 400 µl urine sample to 200 µl phosphate buffer (0.2 M Na2HPO4/0.2 M NaH2PO4, pH 7.4), which was incubated at room temperature for 15 minutes. Each urine sample was then centrifuged for 15 min at 2380 rcf and 500 µl of the supernatant was collected and stored at -80°C overnight. Once thawed, each sample was added to a 5 mm NMR tube with a 50 µl solution of 1mg ml-1 3-(trimethylsilyl)-3,3,2,2-tetradeuteropropionic acid sodium salt (TSP), and 1mg ml-1 sodium azide in deuterium oxide (D2O). TSP was added as an internal reference giving the opportunity for quantitative analysis if required and giving a positional point of reference, as TSP is referenced to 0 ppm. Sodium azide served as a bactericide, and D2O provided the field frequency lock signal for the NMR spectrometer. The protocol for the second set of urine samples provided a more robust method for long term storage and stability compared to the first set of samples. This protocol was used to decrease the chance of degradation of the samples prior to testing. Comparison of the two groups of urine samples showed no degradation or difference in quality, enabling the two groups to be compressed into one thus increasing the power of the statistical analysis.
All samples were analysed using a Bruker AVANCE 600 with a TCI cryo-probe at a proton frequency of 600.17 MHz. Experiments were conducted at 25°C using the pulse sequence for excitation sculpting, as found in the standard Bruker Pulse Library, in order to remove the water signal. The spectra of urine samples were acquired using 64 free induction decays and 16K data points, with a repetition time of 2.5 seconds and a sweep width of 7788 Hz.
Spectra were analysed using the program TOPSPIN (version 1.3) in order to calculate the area under the curves in each transformed spectrum. The area of the allantoin peaks was compared against creatinine as a normalising factor that has been frequently used as a stable normaliser in mice urine [22Rey S, Iturriaga R. Curr Neurovasc Res 2004; 1: 465.
[http://dx.doi.org/10.2174/1567202043361857] [PMID: 16181094] -24Trzebski A, Sato Y, Suzuki A, Sato A. Neurosci Lett 1995; 190: 29.
[http://dx.doi.org/10.1016/0304-3940(95)11492-F] ].
The identification of allantoin through spectral peak analysis used a combination of several two dimensional (2D) experiments to determine the connectivities of 13C and 1H resonances. These experiments included correlation spectro-scopy (COSY), nuclear overhauser spectroscopy (NOESY), total correlation spectroscopy (TOCSY), heteronuclear single quantum coherence (HSQC), and heteronuclear multiple bond coherence (HMBC).
VALIDATION OF PEAKS FROM NMR SPECTRA
Urine samples were spiked with allantoin to validate the 2D analysis. Samples were also spiked with creatinine in order to determine the location of peaks corresponding to this compound for subsequent use as a normaliser for statistical analysis.
Briefly, 4.7 mg of allantoin was dissolved in 200 µl D20, to give a 150 mM solution. This was mixed vigorously and placed in a shaking heating block at 650 rpm and 65°C until allantoin dissolved, and it was stored at room temperature until required. Immediately prior to use, the solution was briefly centrifuged to remove undissolved allantoin that would interfere with NMR analysis. A 50 µl aliquot of the supernatant was added to 500 µl of undiluted mouse urine. This concentration was considered lower than the optimum concentration due to the difficulty in dissolving allantoin; however, positive validation was achieved. Creatinine was prepared using the same method.
STATISTICAL ANALYSIS
A Mann-Whitney test was used to test for case-control differences for the median of the metabolite ratio variable (e.g. allantoin/creatinine) and HIS. The sample correlation between the metabolite ratio variable and the HIS was computed for the set of mdr1a gene targeted mutant mice and the set of control mice. A permutation test was also used to determine the correlation between the HIS and the metabolite ratio score in the set of mdr1a gene targeted mutant mice and the set of control mice. More precisely, for a set of mice (mdr1a gene targeted mutant or control mice), the histological injury scores of the mice were randomly permuted 500,000 times, and for each permutation the sample correlation of the permuted HIS and the unpermuted metabolite ratio score was computed. If k of the permutations yielded a sample correlation greater than or equal to the sample correlation of the ratio score and the unpermuted HIS, then the correlation was significant at the (k+1)/(500,001) level [25Wang ZZ, Stensaas LJ, Bredt DS, Dinger B, Fidone SJ. Neuroscience 1994; 60: 275.
[http://dx.doi.org/10.1016/0306-4522(94)90221-6] ]. Results with p-values less than 0.05 were deemed statistically significant.
RESULTS
The histological injury scores (HIS) of the mice ranged from 0-6 for the control mice and 3-28 for the mdr1a -/- gene targeted mutant mice, averages and standard deviations seen in Fig (2A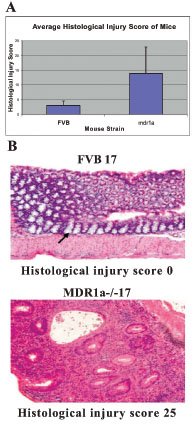 ). The mice contributing to the results described here showed a wide range of disease stages. Fig. (2B) illustrates the phenotypic difference between the inflamed and non-inflamed tissues relating to these disease stages. Fig. (3A
). The mice contributing to the results described here showed a wide range of disease stages. Fig. (2B) illustrates the phenotypic difference between the inflamed and non-inflamed tissues relating to these disease stages. Fig. (3A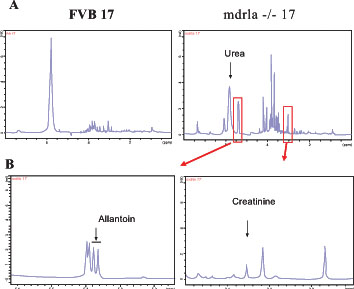 ) shows the difference in NMR profiles of the urine from the same two mice.
) shows the difference in NMR profiles of the urine from the same two mice.


To identify the structure of allantoin, two dimensional analysis was performed on the urine of one homozygous mdr1a gene mutant mouse and one control FVB mouse. A number of two dimensional structural analysis programmes were used to identify the peaks relating to allantoin, which is shown in Fig. (3A). Fig. (4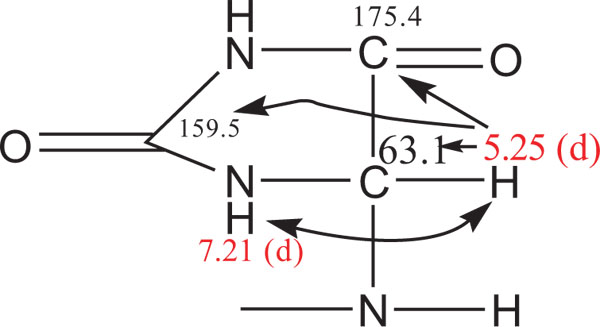 ) provides a summary of the 1H and 13C data that was used to identify the structure of allantoin. Once the allantoin peaks were identified, all 46 urine samples were processed using one dimensional proton (1H) NMR spectroscopy.
) provides a summary of the 1H and 13C data that was used to identify the structure of allantoin. Once the allantoin peaks were identified, all 46 urine samples were processed using one dimensional proton (1H) NMR spectroscopy.
 |
Fig. (4) 1H and 13C resonances of allantoin identified in mouse urine by 2D NMR experiments. |
The integrated area of the allantoin peak was normalised against two internal controls, urea and creatinine; peak locations are shown in Fig. (3). The latter has been used as a relevant normaliser in other studies [22Rey S, Iturriaga R. Curr Neurovasc Res 2004; 1: 465.
[http://dx.doi.org/10.2174/1567202043361857] [PMID: 16181094] -24Trzebski A, Sato Y, Suzuki A, Sato A. Neurosci Lett 1995; 190: 29.
[http://dx.doi.org/10.1016/0304-3940(95)11492-F] , 26Fung ML, Ye JS, Fung PC. Pflugers Arch 2001; 442: 903.
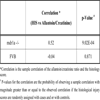
[http://dx.doi.org/10.1007/s004240100610] [PMID: 11680624] ], while urea was used as a further validating control. A Mann-Whitney test showed significant differences in the medians between the mdr1a gene targeted mutant mice (case) and control mice for all metabolite ratio scores and HIS (Table 1). Case-control differences in the median HIS, the allantoin/urea ratio score, and the allantoin/creatinine ratio score were statistically significant (p < 10-6).

We also tested the null hypothesis that the HIS of colonic tissue samples were uncorrelated with the metabolite ratio scores in both the mdr1a gene targeted mutant mice and the control mice. Using creatinine as the internal normaliser, the sample correlation between the metabolite ratio score and the HIS in the mdr1a gene targeted mutant mice is 0.52 as show in Table 2. Permutation testing showed that all correlations were statistically significant at the 0.05 significance level (Table 2). There was no significant correlation between the FVB (control) mice metabolite ratio scores and the HIS at the p=0.05 level (Table 2). Therefore, the allantoin peaks appear to be positively correlated with the HIS in the mdr1a gene targeted mutant mice, but not in the control mice.
DISCUSSION
Allantoin levels have been reported as a sensitive marker of oxidative stress in multiple studies [9Tipoe GL, Fung ML. Respir Physiol Neurobiol 2003; 138: 143.
[http://dx.doi.org/10.1016/S1569-9048(03)00188-5] , 10Milkiewicz M, Pugh CW, Egginton S. J Physiol 2004; 560: 21.
[http://dx.doi.org/10.1113/jphysiol.2004.069757] [PMID: 15319416] [PMCID: PMC1665195] , 27Ye JS, Tipoe GL, Fung PC, Fung ML. Pflugers Arch 2002; 444: 178.
[http://dx.doi.org/10.1007/s00424-002-0785-1] [PMID: 11976930] ]. The data reported here show that allantoin might also represent a novel biomarker for gut inflammation, involving oxidative stress, in the mdr1a -/- gene targeted mutant mouse model.
Allantoin is a product of the oxidation of uric acid by reactive oxygen species (ROS) in humans and hominoid primates [11Roy A, Li J, Baby SM, Mokashi A, Buerk DG, Lahiri S. Respir Physiol Neurobiol 2004; 141: 115.
[http://dx.doi.org/10.1016/j.resp.2004.03.010] [PMID: 15239962] ]. However, allantoin is produced by two independent pathways in the mouse, one involving ROS and the other a reaction between uric acid and urate oxidase (Fig. 1) [28Tjong YW, Jian K, Li M, Chen M, Gao TM, Fung ML. Free Radic Biol Med 2007; 42: 52.
[http://dx.doi.org/10.1016/j.freeradbiomed.2006.09.020] [PMID: 17157193] , 29Elvidge GP, Glenny L, Appelhoff RJ, Ratcliffe PJ, Ragoussis J, Gleadle JM. J Biol Chem 2006; 281: 15215.
[http://dx.doi.org/10.1074/jbc.M511408200] [PMID: 16565084] ]. It is not clear what pathway is predo-minantly responsible for elevated levels of allantoin in mice carrying the mdr1a gene targeted mutation, and further work needs to be undertaken to prove this trend in other CD models, and in human patients.
The work reported here also shows that metabolite profiling through 1H NMR spectroscopy is a time efficient, non-destructive method that requires little preparation of the samples. This method can provide both qualitative and quantitative information of the urinary metabolites with the potential for structural information in 2D spectra if required [15Prasad M, Fearon IM, Zhang M, Laing M, Vollmer C, Nurse CA. J Physiol 2001; 537: 667.
[http://dx.doi.org/10.1113/jphysiol.2001.012836] ]. However, the approach described here concerned the use of internal normalisation to determine relative levels of allantoin. In this respect there appears to be little agreement in the literature on a normalisation method for semi-quantitative analysis of metabolites in urine. This paper has assumed, as have Saude et al. (2007), that each mouse urine sample has a constant level of creatinine and so it was used to reference allantoin peak heights.
Metabolite profiling of urine could be used in human studies to provide a simple and effective means of assessing disease state. As a consequence, food trials that are directed to reducing gut inflammation in susceptible patients could be assessed.
ACKNOWLEDGEMENTS
We acknowledge the financial support of the Foundation for Science, Research and Technology (New Zealand), the Health Research Council of New Zealand, the Marsden Fund, and the University of Auckland Research Committee.
REFERENCES
| [1] | Gonzalez C, Almaraz L, Obeso A, Rigual R. Physiol Rev 1994; 74: 829. [PMID: 7938227] |
| [2] | Gonzalez C, Lopez-Lopez JR, Obeso A, Perez-Garcia MT, Rocher A. Respir Physiol 1995; 102: 137. [http://dx.doi.org/10.1016/0034-5687(95)00069-0] |
| [3] | Riesco-Fagundo AM, Perez-Garcia MT, Gonzalez C, Lopez-Lopez JR. Circ Res 2001; 89: 430. [http://dx.doi.org/10.1161/hh1701.095632] [PMID: 11532904] |
| [4] | Lahiri S, Roy A, Baby SM, Hoshi T, Semenza GL, Prabhakar NR. Prog Biophys Mol Biol 2006; 91: 249. [http://dx.doi.org/10.1016/j.pbiomolbio.2005.07.001] [PMID: 16137743] |
| [5] | Semenza GL. Hydroxylation of HIF-1: oxygen sensing at the molecular level Physiology (Bethesda) 2004; 19: 176. [http://dx.doi.org/10.1152/physiol.00001.2004] [PMID: 15304631] |
| [6] | Semenza GL. J Appl Physiol 2004; 96: 1170. |
| [7] | Baby SM, Roy A, Mokashi AM, Lahiri S. Histochem Cell Biol 2003; 120: 343. [http://dx.doi.org/10.1007/s00418-003-0588-2] [PMID: 14600837] |
| [8] | Roy A, Volgin DV, Baby SM, Mokashi A, Kubin L, Lahiri S. Neurosci Lett 2004; 363: 229. [http://dx.doi.org/10.1016/j.neulet.2004.03.073] [PMID: 15182949] |
| [9] | Tipoe GL, Fung ML. Respir Physiol Neurobiol 2003; 138: 143. [http://dx.doi.org/10.1016/S1569-9048(03)00188-5] |
| [10] | Milkiewicz M, Pugh CW, Egginton S. J Physiol 2004; 560: 21. [http://dx.doi.org/10.1113/jphysiol.2004.069757] [PMID: 15319416] [PMCID: PMC1665195] |
| [11] | Roy A, Li J, Baby SM, Mokashi A, Buerk DG, Lahiri S. Respir Physiol Neurobiol 2004; 141: 115. [http://dx.doi.org/10.1016/j.resp.2004.03.010] [PMID: 15239962] |
| [12] | Vicario I, Rigual R, Obeso A, Gonzalez C. Am J Physiol Cell Physiol 2000; 278: C490. [PMID: 10712237] |
| [13] | Nurse CA. Auton Neurosci 2005; 120: 1. [http://dx.doi.org/10.1016/j.autneu.2005.04.008] [PMID: 15955746] |
| [14] | Lahiri S, Baby SM, Roy A. Curr Respir Med Rev 2006; 2: 393. [http://dx.doi.org/10.2174/157339806778777221] |
| [15] | Prasad M, Fearon IM, Zhang M, Laing M, Vollmer C, Nurse CA. J Physiol 2001; 537: 667. [http://dx.doi.org/10.1113/jphysiol.2001.012836] |
| [16] | Zhang M, Zhong H, Vollmer C, Nurse CA. J Physiol 2000; 525: 143. [http://dx.doi.org/10.1111/j.1469-7793.2000.t01-1-00143.x] [PMID: 11254767 ] [PMCID: PMC2269919] |
| [17] | Grimes PA, Mokashi A, Stone RA, Lahiri S. J Auton Nerv Syst 1995; 54: 80. [http://dx.doi.org/10.1016/0165-1838(95)00006-J] |
| [18] | Iturriaga R, Villanueva S, Mosqueira M. J Appl Physiol 2000; 89: 1005. |
| [19] | Prabhakar NR. Respir Physiol 1999; 115: 161. [http://dx.doi.org/10.1016/S0034-5687(99)00019-5] |
| [20] | Wang ZZ, Dinger BG, Stensaas LJ, Fidone SJ. Biol Signals 1995; 4: 109. [http://dx.doi.org/10.1159/000109430] [PMID: 8750936] |
| [21] | Wang ZZ, Stensaas LJ, Dinger BG, Fidone SJ. Neuroscience 1995; 65: 217. [http://dx.doi.org/10.1016/0306-4522(94)00437-A] |
| [22] | Rey S, Iturriaga R. Curr Neurovasc Res 2004; 1: 465. [http://dx.doi.org/10.2174/1567202043361857] [PMID: 16181094] |
| [23] | Chugh DK, Katayama M, Mokashi A, Bebout DE, Ray DK, Lahiri S. Respir Physiol 1994; 97: 147. [http://dx.doi.org/10.1016/0034-5687(94)90022-1] |
| [24] | Trzebski A, Sato Y, Suzuki A, Sato A. Neurosci Lett 1995; 190: 29. [http://dx.doi.org/10.1016/0304-3940(95)11492-F] |
| [25] | Wang ZZ, Stensaas LJ, Bredt DS, Dinger B, Fidone SJ. Neuroscience 1994; 60: 275. [http://dx.doi.org/10.1016/0306-4522(94)90221-6] |
| [26] | Fung ML, Ye JS, Fung PC. Pflugers Arch 2001; 442: 903. [http://dx.doi.org/10.1007/s004240100610] [PMID: 11680624] |
| [27] | Ye JS, Tipoe GL, Fung PC, Fung ML. Pflugers Arch 2002; 444: 178. [http://dx.doi.org/10.1007/s00424-002-0785-1] [PMID: 11976930] |
| [28] | Tjong YW, Jian K, Li M, Chen M, Gao TM, Fung ML. Free Radic Biol Med 2007; 42: 52. [http://dx.doi.org/10.1016/j.freeradbiomed.2006.09.020] [PMID: 17157193] |
| [29] | Elvidge GP, Glenny L, Appelhoff RJ, Ratcliffe PJ, Ragoussis J, Gleadle JM. J Biol Chem 2006; 281: 15215. [http://dx.doi.org/10.1074/jbc.M511408200] [PMID: 16565084] |
| [30] | Ortega-Saenz P, Garcia-Fernandez M, Pardal R, Alvarez E, Lopez-Barneo J. Adv Exp Med Biol 2003; 536: 65. [http://dx.doi.org/10.1007/978-1-4419-9280-2_9] [PMID: 14635651] |
| [31] | He L, Chen J, Dinger B, Stensaas L, Fidone S. J Appl Physiol 2006; 100: 157. [http://dx.doi.org/10.1152/japplphysiol.00859.2005] [PMID: 16357082] |
| [32] | Valdes V, Mosqueira M, Rey S, Del Rio R, Iturriaga R. Am J Physiol Lung Cell Mol Physiol 2003; 284: L57. [PMID: 12388352] |
| [33] | Wang ZZ, Bredt DS, Fidone SJ, Stensaas LJ. J Comp Neurol 1993; 336: 419. [http://dx.doi.org/10.1002/cne.903360308] [PMID: 7505296] |
| [34] | Prabhakar NR, Kumar GK, Chang CH, Agani FH, Haxhiu MA. Brain Res 1993; 625: 16. [http://dx.doi.org/10.1016/0006-8993(93)90132-7] |
| [35] | Wei G, Dawson VL, Zweier JL. Biochim Biophys Acta 1999; 1455: 23. [http://dx.doi.org/10.1016/S0925-4439(99)00051-4] |
| [36] | Vincent SR. Prog Neurobiol 1994; 42: 129. [http://dx.doi.org/10.1016/0301-0082(94)90023-X] |
| [37] | Vincent SR. J Chem Neuroanat 2000; 18: 23. [http://dx.doi.org/10.1016/S0891-0618(99)00048-4] |
| [38] | Yamamoto Y, Konig P, Henrich M, Dedio J, Kummer W. Cell Tissue Res 2006; 325: 3. [http://dx.doi.org/10.1007/s00441-006-0178-4] [PMID: 16534602] |
| [39] | Campanucci VA, Zhang M, Vollmer C, Nurse CA. J Neurosci 2006; 26: 9482. [http://dx.doi.org/10.1523/JNEUROSCI.1672-06.2006] [PMID: 16971532] |
| [40] | Silva G, Beierwaltes WH, Garvin JL. Hypertension 2006; 47: 563. [http://dx.doi.org/10.1161/01.HYP.0000197954.93874.ef] [PMID: 16380539] |
| [41] | Harrington LS, Mitchell JA. Br J Pharmacol 2004; 143: 611. [http://dx.doi.org/10.1038/sj.bjp.0706004] [PMID: 15466440] [PMCID: PMC1575439] |
| [42] | Summers BA, Overholt JL, Prabhakar NR. J Neurophysiol 1999; 81: 1449. |
| [43] | Postovit LM, Sullivan R, Adams MA, Graham CH. Toxicology 2005; 208: 235. [http://dx.doi.org/10.1016/j.tox.2004.11.022] [PMID: 15691588] |
| [44] | Kasuno K, Takabuchi S, Fukuda K, et al. Biol Chem 2004; 279: 2550. |
| [45] | Berchner-Pfannschmidt U, Yamac H, Trinidad B, Fandrey JJ. Biol Chem 2007; 282: 1788. |