- Home
- About Journals
-
Information for Authors/ReviewersEditorial Policies
Publication Fee
Publication Cycle - Process Flowchart
Online Manuscript Submission and Tracking System
Publishing Ethics and Rectitude
Authorship
Author Benefits
Reviewer Guidelines
Guest Editor Guidelines
Peer Review Workflow
Quick Track Option
Copyediting Services
Bentham Open Membership
Bentham Open Advisory Board
Archiving Policies
Fabricating and Stating False Information
Post Publication Discussions and Corrections
Editorial Management
Advertise With Us
Funding Agencies
Rate List
Kudos
General FAQs
Special Fee Waivers and Discounts
- Contact
- Help
- About Us
- Search




The Open Virology Journal
(Discontinued)
ISSN: 1874-3579 ― Volume 15, 2021
Exosome Biogenesis and Biological Function in Response to Viral Infections
Brennetta J. Crenshaw1, Linlin Gu2, Brian Sims3, Qiana L. Matthews1, *
Abstract
Introduction:
Exosomes are extracellular vesicles that originate as intraluminal vesicles during the process of multivescular body formation. Exosomes mediate intercellular transfer of functional proteins, lipids, and RNAs. The investigation into the formation and role of exosomes in viral infections is still being elucidated. Exosomes and several viruses share similar structural and molecular characteristics.
Explanation:
It has been documented that viral hijacking exploits the exosomal pathway and mimics cellular protein trafficking. Exosomes released from virus-infected cells contain a variety of viral and host cellular factors that are able to modify recipient host cell responses. Recent studies have demonstrated that exosomes are crucial components in the pathogenesis of virus infection. Exosomes also allow the host to produce effective immunity against pathogens by activating antiviral mechanisms and transporting antiviral factors between adjacent cells.
Conclusion:
Given the ever-growing roles and importance of exosomes in both host and pathogen response, this review will address the impact role of exosome biogenesis and composition after DNA, RNA virus, on Retrovirus infections. This review also will also address how exosomes can be used as therapeutic agents as well as a vaccine vehicles.
Article Information

Identifiers and Pagination:
Year: 2018Volume: 12
First Page: 134
Last Page: 148
Publisher Id: TOVJ-12-134
DOI: 10.2174/1874357901812010134
Article History:
Received Date: 1/2/2018Revision Received Date: 27/5/2018
Acceptance Date: 8/8/2018
Electronic publication date: 28/09/2018
Collection year: 2018
open-access license: This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International Public License (CC-BY 4.0), a copy of which is available at: https://creativecommons.org/licenses/by/4.0/legalcode. This license permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
* Address correspondence to this author at the Department of Biological Sciences, Microbiology Program, College of Science, Technology, Engineering and Mathematics, Alabama State University, Montgomery, 36104 AL, USA; Tel: 334-229-8449; E-mail:qmatthews@alasu.edu
| Open Peer Review Details | |||
|---|---|---|---|
| Manuscript submitted on 1-2-2018 |
Original Manuscript | Exosome Biogenesis and Biological Function in Response to Viral Infections | |
1. INTRODUCTION
Exosomes are minute (30-150nm) extracellular vesicles (EVs) that are generated during the maturation of endosomes [1Chahar HS, Bao X, Casola A. Exosomes and their role in the life cycle and pathogenesis of RNA viruses. Viruses 2015; 7(6): 3204-25.
[http://dx.doi.org/10.3390/v7062770] [PMID: 26102580] ]. Exosomes are secreted into the extracellular environment by several cell types, such as tumor and immune cells [2Théry C. Exosomes: Secreted vesicles and intercellular communications. F1000 Biol Rep 2011; 3: 15.
[http://dx.doi.org/10.3410/B3-15] [PMID: 21876726] ], antigen-presenting cells [3Raposo G, Stoorvogel W. Extracellular vesicles: Exosomes, microvesicles, and friends. J Cell Biol 2013; 200(4): 373-83.
[http://dx.doi.org/10.1083/jcb.201211138] [PMID: 23420871] ], and epithelial cells [4van Niel G, Raposo G, Candalh C, et al. Intestinal epithelial cells secrete exosome-like vesicles. Gastroenterology 2001; 121(2): 337-49.
[http://dx.doi.org/10.1053/gast.2001.26263] [PMID: 11487543] ]. Exosomes are found in biological fluids such as blood [5Thery C, Amigorena S, Raposo G, et al. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr Protoc Cell Biol 2006. Chapter 3; Unit 3 22.
[http://dx.doi.org/10.1002/0471143030.cb0322s30] [PMID: 18228490] ], urine [6De Toro J, Herschlik L, Waldner C, Mongini C. Emerging roles of exosomes in normal and pathological conditions: new insights for diagnosis and therapeutic applications. Front Immunolo 2015; 6(203): 1-12.
[http://dx.doi.org/10.3389/fimmu.2015.00203] [PMID: 25999947] , 7Dear JW, Street JM, Bailey MA. Urinary exosomes: A reservoir for biomarker discovery and potential mediators of intrarenal signalling. Proteomics 2013; 13(10-11): 1572-80.
[http://dx.doi.org/10.1002/pmic.201200285] [PMID: 23129434] ], semen [6De Toro J, Herschlik L, Waldner C, Mongini C. Emerging roles of exosomes in normal and pathological conditions: new insights for diagnosis and therapeutic applications. Front Immunolo 2015; 6(203): 1-12.
[http://dx.doi.org/10.3389/fimmu.2015.00203] [PMID: 25999947] , 8Madison MN, Roller RJ, Okeoma CM. Human semen contains exosomes with potent anti-HIV-1 activity. Retrovirology 2014; 11: 102.
[http://dx.doi.org/10.1186/s12977-014-0102-z] [PMID: 25407601] ], saliva [6De Toro J, Herschlik L, Waldner C, Mongini C. Emerging roles of exosomes in normal and pathological conditions: new insights for diagnosis and therapeutic applications. Front Immunolo 2015; 6(203): 1-12.
[http://dx.doi.org/10.3389/fimmu.2015.00203] [PMID: 25999947] , 9Lässer C, Alikhani VS, Ekström K, et al. Human saliva, plasma and breast milk exosomes contain RNA: Uptake by macrophages. J Transl Med 2011; 9: 9.
[http://dx.doi.org/10.1186/1479-5876-9-9] [PMID: 21235781] ], cerebrospinal fluid [6De Toro J, Herschlik L, Waldner C, Mongini C. Emerging roles of exosomes in normal and pathological conditions: new insights for diagnosis and therapeutic applications. Front Immunolo 2015; 6(203): 1-12.
[http://dx.doi.org/10.3389/fimmu.2015.00203] [PMID: 25999947] , 10Street JM, Barran PE, Mackay CL, et al. Identification and proteomic profiling of exosomes in human cerebrospinal fluid. J Transl Med 2012; 10: 5.
[http://dx.doi.org/10.1186/1479-5876-10-5] [PMID: 22221959] ] and breast milk [6De Toro J, Herschlik L, Waldner C, Mongini C. Emerging roles of exosomes in normal and pathological conditions: new insights for diagnosis and therapeutic applications. Front Immunolo 2015; 6(203): 1-12.
[http://dx.doi.org/10.3389/fimmu.2015.00203] [PMID: 25999947] , 11Näslund TI, Paquin-Proulx D, Paredes PT, Vallhov H, Sandberg JK, Gabrielsson S. Exosomes from breast milk inhibit HIV-1 infection of dendritic cells and subsequent viral transfer to CD4+ T cells. AIDS 2014; 28(2): 171-80.
[http://dx.doi.org/10.1097/QAD.0000000000000159] [PMID: 24413309] ]. Recently, exosomes have developed as an essential tool for facilitating intercellular communication through the transfer of biologically active RNAs, lipids, and proteins [1Chahar HS, Bao X, Casola A. Exosomes and their role in the life cycle and pathogenesis of RNA viruses. Viruses 2015; 7(6): 3204-25.
[http://dx.doi.org/10.3390/v7062770] [PMID: 26102580] , 12Meckes DG Jr, Raab-Traub N. Microvesicles and viral infection. J Virol 2011; 85(24): 12844-54.
[http://dx.doi.org/10.1128/JVI.05853-11] [PMID: 21976651] ].
Exosomes were first observed in the early 1980s by two groups studying the culture media of reticulocytes [13Colombo M, Moita C, van Niel G, et al. Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. J Cell Sci 2013; 126(Pt 24): 5553-65.
[http://dx.doi.org/10.1242/jcs.128868] [PMID: 24105262] -15Pan BT, Johnstone RM. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: Selective externalization of the receptor. Cell 1983; 33(3): 967-78.
[http://dx.doi.org/10.1016/0092-8674(83)90040-5] [PMID: 6307529] ]. They revealed that small vesicles developed via inward budding inside of an intracellular endosome [13Colombo M, Moita C, van Niel G, et al. Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. J Cell Sci 2013; 126(Pt 24): 5553-65.
[http://dx.doi.org/10.1242/jcs.128868] [PMID: 24105262] -15Pan BT, Johnstone RM. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: Selective externalization of the receptor. Cell 1983; 33(3): 967-78.
[http://dx.doi.org/10.1016/0092-8674(83)90040-5] [PMID: 6307529] ]. In the late 80s, the word “exosomes” was coined by Dr. Rose Johnstone [16Akers JC, Gonda D, Kim R, Carter BS, Chen CC. Biogenesis of Extracellular Vesicles (EV): Exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J Neurooncol 2013; 113(1): 1-11.
[http://dx.doi.org/10.1007/s11060-013-1084-8] [PMID: 23456661] , 17Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J Biol Chem 1987; 262(19): 9412-20.
[PMID: 3597417] ]. She observed that developing reticulocytes are composed of large sacs filled with miniature membrane-enclosed vesicles that were 30-100 nm [16Akers JC, Gonda D, Kim R, Carter BS, Chen CC. Biogenesis of Extracellular Vesicles (EV): Exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J Neurooncol 2013; 113(1): 1-11.
[http://dx.doi.org/10.1007/s11060-013-1084-8] [PMID: 23456661] , 17Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J Biol Chem 1987; 262(19): 9412-20.
[PMID: 3597417] ]. Later, she identified transferrin as a plentiful membrane protein on these sacs [16Akers JC, Gonda D, Kim R, Carter BS, Chen CC. Biogenesis of Extracellular Vesicles (EV): Exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J Neurooncol 2013; 113(1): 1-11.
[http://dx.doi.org/10.1007/s11060-013-1084-8] [PMID: 23456661] , 17Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J Biol Chem 1987; 262(19): 9412-20.
[PMID: 3597417] ]. Immunogold labeling that contained a monoclonal antibody alongside the transferrin receptor showed that the larger sacs joined with the cell’s plasma membrane, secreting the smaller membrane-enclosed structures [16Akers JC, Gonda D, Kim R, Carter BS, Chen CC. Biogenesis of Extracellular Vesicles (EV): Exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J Neurooncol 2013; 113(1): 1-11.
[http://dx.doi.org/10.1007/s11060-013-1084-8] [PMID: 23456661] , 17Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J Biol Chem 1987; 262(19): 9412-20.
[PMID: 3597417] ].
Since their discovery, the biogenesis, secretion, and composition of exosomes have been extensively studied [18Kalra H, Drummen GP, Mathivanan S. Focus on extracellular vesicles: Introducing the next small big thing. Int J Mol Sci 2016; 17(2): 170.
[http://dx.doi.org/10.3390/ijms17020170] [PMID: 26861301] ]. When discovered more than 30 years ago, exosomes were originally thought to be a mechanism of discarding plasma membrane proteins in maturing reticulocytes found in red blood cells [13Colombo M, Moita C, van Niel G, et al. Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. J Cell Sci 2013; 126(Pt 24): 5553-65.
[http://dx.doi.org/10.1242/jcs.128868] [PMID: 24105262] , 18Kalra H, Drummen GP, Mathivanan S. Focus on extracellular vesicles: Introducing the next small big thing. Int J Mol Sci 2016; 17(2): 170.
[http://dx.doi.org/10.3390/ijms17020170] [PMID: 26861301] ]. These small membranous vesicles are generated by the inward budding of the plasma membrane to form intracellular endosomes [19Beach A, Zhang HG, Ratajczak MZ, Kakar SS. Exosomes: An overview of biogenesis, composition and role in ovarian cancer. J Ovarian Res 2014; 7: 14.
[http://dx.doi.org/10.1186/1757-2215-7-14] [PMID: 24460816] ]. Early endosomes join with endocytic vesicles and fuse their content with those intended for exocytosis, degradation, or recycling [16Akers JC, Gonda D, Kim R, Carter BS, Chen CC. Biogenesis of Extracellular Vesicles (EV): Exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J Neurooncol 2013; 113(1): 1-11.
[http://dx.doi.org/10.1007/s11060-013-1084-8] [PMID: 23456661] ]. As the early endosomes mature, they develop into late endosomes and become characterized by the formation of Intraluminal Vesicles (ILV) or Multivesicular Bodies (MVBs) inside the lumen of the endosome [19Beach A, Zhang HG, Ratajczak MZ, Kakar SS. Exosomes: An overview of biogenesis, composition and role in ovarian cancer. J Ovarian Res 2014; 7: 14.
[http://dx.doi.org/10.1186/1757-2215-7-14] [PMID: 24460816] ]. The MVBs fuse with the plasma membrane, as well as lysosomes for degradation of their contents, releasing their contents into the extracellular environment in the form of exosomes [19Beach A, Zhang HG, Ratajczak MZ, Kakar SS. Exosomes: An overview of biogenesis, composition and role in ovarian cancer. J Ovarian Res 2014; 7: 14.
[http://dx.doi.org/10.1186/1757-2215-7-14] [PMID: 24460816] ] (Table 1).

The processes that control the formation of ILVs inside MVBs and the fusion with the plasma membrane to release exosomes into the extracellular environment are not completely understood [18Kalra H, Drummen GP, Mathivanan S. Focus on extracellular vesicles: Introducing the next small big thing. Int J Mol Sci 2016; 17(2): 170.
[http://dx.doi.org/10.3390/ijms17020170] [PMID: 26861301] ]. The biogenesis and secretion of exosomes are believed to be facilitated through the Endosomal Sorting Complex Required for Transport (ESCRT)-dependent pathway [13Colombo M, Moita C, van Niel G, et al. Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. J Cell Sci 2013; 126(Pt 24): 5553-65.
[http://dx.doi.org/10.1242/jcs.128868] [PMID: 24105262] , 18Kalra H, Drummen GP, Mathivanan S. Focus on extracellular vesicles: Introducing the next small big thing. Int J Mol Sci 2016; 17(2): 170.
[http://dx.doi.org/10.3390/ijms17020170] [PMID: 26861301] ] or ceramide-dependent pathway [18Kalra H, Drummen GP, Mathivanan S. Focus on extracellular vesicles: Introducing the next small big thing. Int J Mol Sci 2016; 17(2): 170.
[http://dx.doi.org/10.3390/ijms17020170] [PMID: 26861301] , 20Trajkovic K, Hsu C, Chiantia S, et al. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science 2008; 319(5867): 1244-7.
[http://dx.doi.org/10.1126/science.1153124] [PMID: 18309083] ]. The ESCRT machinery is composed of four components: ESCRT-0, I, II, and III [18Kalra H, Drummen GP, Mathivanan S. Focus on extracellular vesicles: Introducing the next small big thing. Int J Mol Sci 2016; 17(2): 170.
[http://dx.doi.org/10.3390/ijms17020170] [PMID: 26861301] , 21Bassereau P. Division of labour in ESCRT complexes. Nat Cell Biol 2010; 12(5): 422-3.
[http://dx.doi.org/10.1038/ncb0510-422] [PMID: 20442701] ]. In conjunction with a variety of proteins, the ESCRT machinery is primarily involved in the binding, sorting, and clustering of ubiquitinylated receptors and proteins in the late endosome [18Kalra H, Drummen GP, Mathivanan S. Focus on extracellular vesicles: Introducing the next small big thing. Int J Mol Sci 2016; 17(2): 170.
[http://dx.doi.org/10.3390/ijms17020170] [PMID: 26861301] ]. In the ESCRT-dependent pathway, components of the ESCRT machinery are consecutively transported to the endosomal membrane [18Kalra H, Drummen GP, Mathivanan S. Focus on extracellular vesicles: Introducing the next small big thing. Int J Mol Sci 2016; 17(2): 170.
[http://dx.doi.org/10.3390/ijms17020170] [PMID: 26861301] ]. Along with transportation, they begin with the hepatocyte growth factor-regulated lyrosine kinase substrate (Hrs) and bind to the ubiquitinate protein ESCRT-0, the phosphatidylinositol-3-phosphate (PI(3)P), and 3,5-bisphosphate (PI(3,5)P2) through lipid-bind domains: GRAM-Like Ubiquitin-binding in EAP45 (GLUE) [18Kalra H, Drummen GP, Mathivanan S. Focus on extracellular vesicles: Introducing the next small big thing. Int J Mol Sci 2016; 17(2): 170.
[http://dx.doi.org/10.3390/ijms17020170] [PMID: 26861301] , 22Teo H, Gill DJ, Sun J, et al. ESCRT-I core and ESCRT-II GLUE domain structures reveal role for GLUE in linking to ESCRT-I and membranes. Cell 2006; 125(1): 99-111.
[http://dx.doi.org/10.1016/j.cell.2006.01.047] [PMID: 16615893] ] and Fab-1, YGL023, Vps27, and EEA1 (FYVE) [18Kalra H, Drummen GP, Mathivanan S. Focus on extracellular vesicles: Introducing the next small big thing. Int J Mol Sci 2016; 17(2): 170.
[http://dx.doi.org/10.3390/ijms17020170] [PMID: 26861301] , 23Stahelin RV, Long F, Diraviyam K. Phosphatidylinositol 3-phosphate induces the membrane penetration of the FYVE domains of Vps27p and Hr. J Biol Chem 2002; 277(29): 26379-88.
[http://dx.doi.org/10.1074/jbc.M201106200] [PMID: 12006563] ]. ESCRT-I and -II facilitate the budding of ILVs, in which cargo is transported into the lumen [18Kalra H, Drummen GP, Mathivanan S. Focus on extracellular vesicles: Introducing the next small big thing. Int J Mol Sci 2016; 17(2): 170.
[http://dx.doi.org/10.3390/ijms17020170] [PMID: 26861301] ]. ESCRT-III is recruited by ALG-2 interacting protein X (Alix) to facilitate pulling, spiral formation, and complete budding [18Kalra H, Drummen GP, Mathivanan S. Focus on extracellular vesicles: Introducing the next small big thing. Int J Mol Sci 2016; 17(2): 170.
[http://dx.doi.org/10.3390/ijms17020170] [PMID: 26861301] ]. Deubiquitinatings enzyme (DUB) deubiquitinate the protein and vacuolar protein sorting proteins (Vps) 4 reprocess the ESCRT machinery [18Kalra H, Drummen GP, Mathivanan S. Focus on extracellular vesicles: Introducing the next small big thing. Int J Mol Sci 2016; 17(2): 170.
[http://dx.doi.org/10.3390/ijms17020170] [PMID: 26861301] ]. Next, the MVB is transported to the plasma membrane [18Kalra H, Drummen GP, Mathivanan S. Focus on extracellular vesicles: Introducing the next small big thing. Int J Mol Sci 2016; 17(2): 170.
[http://dx.doi.org/10.3390/ijms17020170] [PMID: 26861301] ]. Through fusion, the ILVs are released into the extracellular environment and are denoted as “exosomes” [18Kalra H, Drummen GP, Mathivanan S. Focus on extracellular vesicles: Introducing the next small big thing. Int J Mol Sci 2016; 17(2): 170.
[http://dx.doi.org/10.3390/ijms17020170] [PMID: 26861301] ].
In addition to the ESCRT-dependent pathway, current research has reported the presence of an alternate pathway referred to as the ceramide-dependent pathway [18Kalra H, Drummen GP, Mathivanan S. Focus on extracellular vesicles: Introducing the next small big thing. Int J Mol Sci 2016; 17(2): 170.
[http://dx.doi.org/10.3390/ijms17020170] [PMID: 26861301] , 20Trajkovic K, Hsu C, Chiantia S, et al. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science 2008; 319(5867): 1244-7.
[http://dx.doi.org/10.1126/science.1153124] [PMID: 18309083] , 24Stuffers S, Sem Wegner C, Stenmark H, Brech A. Multivesicular endosome biogenesis in the absence of ESCRTs. Traffic 2009; 10(7): 925-37.
[http://dx.doi.org/10.1111/j.1600-0854.2009.00920.x] [PMID: 19490536] ]. The ceramide-dependent pathway is based on the development of glycolipoprotein microdomains (lipid rafts) in which sphingomyelin is transformed into ceramide by sphingomyelinases (enzymes generating ceramide from sphingomyelin) [18Kalra H, Drummen GP, Mathivanan S. Focus on extracellular vesicles: Introducing the next small big thing. Int J Mol Sci 2016; 17(2): 170.
[http://dx.doi.org/10.3390/ijms17020170] [PMID: 26861301] , 20Trajkovic K, Hsu C, Chiantia S, et al. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science 2008; 319(5867): 1244-7.
[http://dx.doi.org/10.1126/science.1153124] [PMID: 18309083] , 24Stuffers S, Sem Wegner C, Stenmark H, Brech A. Multivesicular endosome biogenesis in the absence of ESCRTs. Traffic 2009; 10(7): 925-37.
[http://dx.doi.org/10.1111/j.1600-0854.2009.00920.x] [PMID: 19490536] ]. Ceramide accumulation then prompts microdomain amalgamation and initiates ILV formation within MVBs [18Kalra H, Drummen GP, Mathivanan S. Focus on extracellular vesicles: Introducing the next small big thing. Int J Mol Sci 2016; 17(2): 170.
[http://dx.doi.org/10.3390/ijms17020170] [PMID: 26861301] ].
Exosomes are secreted in many cell types during normal, physiological, and pathological conditions [6De Toro J, Herschlik L, Waldner C, Mongini C. Emerging roles of exosomes in normal and pathological conditions: new insights for diagnosis and therapeutic applications. Front Immunolo 2015; 6(203): 1-12.
[http://dx.doi.org/10.3389/fimmu.2015.00203] [PMID: 25999947] , 25Yuana Y, Sturk A, Nieuwland R. Extracellular vesicles in physiological and pathological conditions. Blood Rev 2013; 27(1): 31-9.
[http://dx.doi.org/10.1016/j.blre.2012.12.002] [PMID: 23261067] , 26Beach A, Zhang HG, Ratajczak MZ, Kakar SS. Exosomes: An overview of biogenesis, composition and role in ovarian cancer. J Ovarian Res 2014; 7: 14-4.
[http://dx.doi.org/10.1186/1757-2215-7-14] [PMID: 24460816] ]. The regulatory molecules involving the release of exosomes were reported by Ostrowski et al. [27Zhang J, Li S, Li L, et al. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genomics Proteomics Bioinformatics 2015; 13(1): 17-24.
[http://dx.doi.org/10.1016/j.gpb.2015.02.001] [PMID: 25724326] , 28Ostrowski M, Carmo NB, Krumeich S, et al. Rab27a and Rab27b control different steps of the exosome secretion pathway. Nat Cell Biol 2010; 12(1): 19-30.
[http://dx.doi.org/10.1038/ncb2000] [PMID: 19966785] ]. They reported that Rab27a and Rab27b were affiliated with exosome secretion [27Zhang J, Li S, Li L, et al. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genomics Proteomics Bioinformatics 2015; 13(1): 17-24.
[http://dx.doi.org/10.1016/j.gpb.2015.02.001] [PMID: 25724326] , 28Ostrowski M, Carmo NB, Krumeich S, et al. Rab27a and Rab27b control different steps of the exosome secretion pathway. Nat Cell Biol 2010; 12(1): 19-30.
[http://dx.doi.org/10.1038/ncb2000] [PMID: 19966785] ]. They also reported that knockdown of Rab27 and/or their effectors, EXPH5, and SYTL4, could prevent the secretion of exosomes in HeLa cells [27Zhang J, Li S, Li L, et al. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genomics Proteomics Bioinformatics 2015; 13(1): 17-24.
[http://dx.doi.org/10.1016/j.gpb.2015.02.001] [PMID: 25724326] , 28Ostrowski M, Carmo NB, Krumeich S, et al. Rab27a and Rab27b control different steps of the exosome secretion pathway. Nat Cell Biol 2010; 12(1): 19-30.
[http://dx.doi.org/10.1038/ncb2000] [PMID: 19966785] ]. Another study was reported by Baietti et al. [27Zhang J, Li S, Li L, et al. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genomics Proteomics Bioinformatics 2015; 13(1): 17-24.
[http://dx.doi.org/10.1016/j.gpb.2015.02.001] [PMID: 25724326] , 29Baietti MF, Zhang Z, Mortier E, et al. Syndecan-syntenin-ALIX regulates the biogenesis of exosomes. Nat Cell Biol 2012; 14(7): 677-85.
[http://dx.doi.org/10.1038/ncb2502] [PMID: 22660413] ]. They reported that syndecan-syntenin networked with ALIX protein through Leu-Tyr-Pro-X(n)-Leu motif to support the intraluminal budding of endosomal membranes [27Zhang J, Li S, Li L, et al. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genomics Proteomics Bioinformatics 2015; 13(1): 17-24.
[http://dx.doi.org/10.1016/j.gpb.2015.02.001] [PMID: 25724326] , 29Baietti MF, Zhang Z, Mortier E, et al. Syndecan-syntenin-ALIX regulates the biogenesis of exosomes. Nat Cell Biol 2012; 14(7): 677-85.
[http://dx.doi.org/10.1038/ncb2502] [PMID: 22660413] ]. Moreover, Yu and colleagues revealed that the tumor repressor protein p53 and its downstream effector TSAP6 could augment exosome production [27Zhang J, Li S, Li L, et al. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genomics Proteomics Bioinformatics 2015; 13(1): 17-24.
[http://dx.doi.org/10.1016/j.gpb.2015.02.001] [PMID: 25724326] , 30Yu X, Harris SL, Levine AJ. The regulation of exosome secretion: A novel function of the p53 protein. Cancer Res 2006; 66(9): 4795-801.
[http://dx.doi.org/10.1158/0008-5472.CAN-05-4579] [PMID: 16651434] ]. Overall, these findings suggest that molecules are responsible for the formation and secretion of exosomes in parent cells; and can act as a regulatory network [27Zhang J, Li S, Li L, et al. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genomics Proteomics Bioinformatics 2015; 13(1): 17-24.
[http://dx.doi.org/10.1016/j.gpb.2015.02.001] [PMID: 25724326] ].
Exosomes have been purified and isolated from in vitro cell cultures and biological fluids using several techniques [31Greening DW, Xu R, Ji H, Tauro BJ, Simpson RJ. A protocol for exosome isolation and characterization: Evaluation of ultracentrifugation, density-gradient separation, and immunoaffinity capture methods. Methods Mol Biol 2015; 1295: 179-209.
[http://dx.doi.org/10.1007/978-1-4939-2550-6_15] [PMID: 25820723] ]. The most common method used to isolate exosomes from cell culture supernatants involves a series of ultracentrifugation to eliminate cells and excessive debris [31Greening DW, Xu R, Ji H, Tauro BJ, Simpson RJ. A protocol for exosome isolation and characterization: Evaluation of ultracentrifugation, density-gradient separation, and immunoaffinity capture methods. Methods Mol Biol 2015; 1295: 179-209.
[http://dx.doi.org/10.1007/978-1-4939-2550-6_15] [PMID: 25820723] ]. A series of ultra-centrifugation is performed to pellet the exosomes. The pellet is washed in a vast amount of phosphate buffer saline (PBS) to remove contaminating proteins. The pellet is then centrifuged an additional time at an equivalent high speed. The size of exosomes is equal to that of the ILVs of the MVBs from where they are derived [3Raposo G, Stoorvogel W. Extracellular vesicles: Exosomes, microvesicles, and friends. J Cell Biol 2013; 200(4): 373-83.
[http://dx.doi.org/10.1083/jcb.201211138] [PMID: 23420871] ]. Additional isolation can be accomplished by polymer-based precipitation [32Alvarez ML, Khosroheidari M, Kanchi Ravi R, DiStefano JK. Comparison of protein, microRNA, and mRNA yields using different methods of urinary exosome isolation for the discovery of kidney disease biomarkers. Kidney Int 2012; 82(9): 1024-32.
[http://dx.doi.org/10.1038/ki.2012.256] [PMID: 22785172] ], immuno-affinity capture [33Tauro BJ, Greening DW, Mathias RA, et al. Comparison of ultracentrifugation, density gradient separation, and immunoaffinity capture methods for isolating human colon cancer cell line LIM1863-derived exosomes. Methods 2012; 56(2): 293-304.
[http://dx.doi.org/10.1016/j.ymeth.2012.01.002] [PMID: 22285593] ], size exclusion chromatography [34Li P, Kaslan M, Lee SH, Yao J, Gao Z. Progress in exosome isolation techniques. Theranostics 2017; 7(3): 789-804.
[http://dx.doi.org/10.7150/thno.18133] [PMID: 28255367] ], microfluidics [35Chen C, Skog J, Hsu CH, et al. Microfluidic isolation and transcriptome analysis of serum microvesicles. Lab Chip 2010; 10(4): 505-11.
[http://dx.doi.org/10.1039/B916199F] [PMID: 20126692] ], and ultrafiltration [34Li P, Kaslan M, Lee SH, Yao J, Gao Z. Progress in exosome isolation techniques. Theranostics 2017; 7(3): 789-804.
[http://dx.doi.org/10.7150/thno.18133] [PMID: 28255367] ]. Due to the rising interest in exosomes for therapeutics, commercially available kits (Exosome Isolation kit, Exoquick, and Exo-spin) [3Raposo G, Stoorvogel W. Extracellular vesicles: Exosomes, microvesicles, and friends. J Cell Biol 2013; 200(4): 373-83.
[http://dx.doi.org/10.1083/jcb.201211138] [PMID: 23420871] ] are available to isolate exosomes.
The assessment of the purity of exosomal preparations, in addition to their composition, is often difficult based on the cell type/origin from which the exosomes are derived [5Thery C, Amigorena S, Raposo G, et al. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr Protoc Cell Biol 2006. Chapter 3; Unit 3 22.
[http://dx.doi.org/10.1002/0471143030.cb0322s30] [PMID: 18228490] ]. Furthermore, it is known that there is a relationship between exosomal trafficking and viral hijacking [36Zhang Y, Bergelson JM. Adenovirus receptors. J Virol 2005; 79(19): 12125-31.
[http://dx.doi.org/10.1128/JVI.79.19.12125-12131.2005] [PMID: 16160140] ]. Therefore, when isolating exosomes from virus-infected cells, it is imperative to ensure that the collected material only contains exosomes and no viral particles. Konadu et al. Optimized many exosome characterization techniques [1Chahar HS, Bao X, Casola A. Exosomes and their role in the life cycle and pathogenesis of RNA viruses. Viruses 2015; 7(6): 3204-25.
[http://dx.doi.org/10.3390/v7062770] [PMID: 26102580] ]. These techniques included using electron microscopy to visualize exosomes, examining exosomal protein markers (Annexin V, CD63, CD81, etc.) through immunoblot analysis, and performing nanoparticle tracking (NanoSight) analysis to measure and analyze the size of exosomes [1Chahar HS, Bao X, Casola A. Exosomes and their role in the life cycle and pathogenesis of RNA viruses. Viruses 2015; 7(6): 3204-25.
[http://dx.doi.org/10.3390/v7062770] [PMID: 26102580] , 37Konadu KA, Huang MB, Roth W, et al. Isolation of exosomes from the plasma of HIV-1 positive individuals. J Vis Exp 2016; 107: 53495.
[http://dx.doi.org/10.3791/53495] [PMID: 26780239] ]. Because of their tiny size, exosomes cannot be identified by using standard flow cytometry. Nonetheless, the existence of epitopes on the surface of exosomes can be used to sort/purify exosomes [38Koliha N, Wiencek Y, Heider U, et al. A novel multiplex bead-based platform highlights the diversity of extracellular vesicles. J Extracell Vesicles 2016; 5: 29975.
[http://dx.doi.org/10.3402/jev.v5.29975] [PMID: 26901056] ].
Recent studies have acknowledged exosomes as vital components in viral pathogenesis and immunity [1Chahar HS, Bao X, Casola A. Exosomes and their role in the life cycle and pathogenesis of RNA viruses. Viruses 2015; 7(6): 3204-25.
[http://dx.doi.org/10.3390/v7062770] [PMID: 26102580] ]. Exosomes allow the host to mount effective immune responses against pathogens, which includes activating antiviral mechanisms and transferring antiviral elements among a variety of cells [39Madison MN, Jones PH, Okeoma CM. Exosomes in human semen restrict HIV-1 transmission by vaginal cells and block intravaginal replication of LP-BM5 murine AIDS virus complex. Virology 2015; 482: 189-201.
[http://dx.doi.org/10.1016/j.virol.2015.03.040] [PMID: 25880110] ]. Depending on the nature of the target and pathogen, exosomes can increase or restrict an infection [1Chahar HS, Bao X, Casola A. Exosomes and their role in the life cycle and pathogenesis of RNA viruses. Viruses 2015; 7(6): 3204-25.
[http://dx.doi.org/10.3390/v7062770] [PMID: 26102580] ]. Exosomes that are composed of viral genomes can stimulate viral spread by entering susceptible cells while eluding immune recognition [40van Dongen HM, Masoumi N, Witwer KW, Pegtel DM. Extracellular vesicles exploit viral entry routes for cargo delivery. Microbiol Mol Biol Rev 2016; 80(2): 369-86.
[http://dx.doi.org/10.1128/MMBR.00063-15] [PMID: 26935137] ]. In certain cases, exosomes that contain nucleic acids or viral proteins initiate immune responses in myeloid cells [41Longatti A. The dual role of exosomes in hepatitis A and C virus transmission and viral immune activation. Viruses 2015; 7(12): 6707-15.
[http://dx.doi.org/10.3390/v7122967] [PMID: 26694453] ]. Research on Dengue Virus, human T-cell lymphotropic virus (HTLV), Hepatitis C Virus (HCV), and Human Immunodeficiency Virus (HIV) has proved that exosomes distributed from infected cells transport various regulatory factors, (as reviewed in (Chahar 2015)). These factors include host functional genetic elements to neighboring cells, cellular and viral miRNA, proteins, and viral RNA which help aid in regulating cellular responses and producing infections [1Chahar HS, Bao X, Casola A. Exosomes and their role in the life cycle and pathogenesis of RNA viruses. Viruses 2015; 7(6): 3204-25.
[http://dx.doi.org/10.3390/v7062770] [PMID: 26102580] ]. This review will summarize the biogenesis of exosomes and their role in biological functions in response to viral infections.
2. HIJACKING BY VIRUSES EXPLOITS THE EXOSOMAL PATHWAY
Viruses take control of and exploit cellular replication mechanisms to replicate. When successful, most viruses end up killing their host cell. Some viruses can hijack affiliates of vesicular trafficking and through a sequence of complexes known as ESCRT. Also, they can assimilate viral constituents into exosomes. Viral antigens in exosomes maximize persistence by hiding viral genomes, entrapping the immune system, and maximizing viral infection in uncontaminated cells. Exosomes can be used to present a source of viral antigens that can be targeted for therapeutic uses. Also, exosomes can be used as a biomarker for disease in the regulation and obliteration of some illnesses [42Anderson MR, Kashanchi F, Jacobson S. Exosomes in viral disease. Neurotherapeutics 2016; 13(3): 535-46.
[http://dx.doi.org/10.1007/s13311-016-0450-6] [PMID: 27324390] ].
Several viruses enter cells through the endocytic pathway. Viruses that enter through endocytosis can hijack and use exosomal pathways for their own benefit. Infectious diseases, such as HCV, Zika virus (ZV), West Nile virus (WNV), and DENV enter this pathway by clathrin-mediated or receptor-mediated endocytosis [43Ramakrishnaiah V, Thumann C, Fofana I, et al. Exosome-mediated transmission of hepatitis C virus between human hepatoma Huh7.5 cells. Proc Natl Acad Sci USA 2013; 110(32): 13109-13.
[http://dx.doi.org/10.1073/pnas.1221899110] [PMID: 23878230] -46Hamel R, Dejarnac O, Wichit S, et al. Biology of Zika virus infection in human skin cells. J Virol 2015; 89(17): 8880-96.
[http://dx.doi.org/10.1128/JVI.00354-15] [PMID: 26085147] ]. After “back-fusion” of ILVs, these infectious viruses antagonize late endosomes, thus resulting in the discarding of the viral genome inside of the cytoplasm [44Nour AM, Modis Y. Endosomal vesicles as vehicles for viral genomes. Trends Cell Biol 2014; 24(8): 449-54.
[http://dx.doi.org/10.1016/j.tcb.2014.03.006] [PMID: 24746011] ]. In the case of HCV, the viral genome can linger in ILVs and be secreted inside of the exosomes, where they can perform as infectious particles [41Longatti A. The dual role of exosomes in hepatitis A and C virus transmission and viral immune activation. Viruses 2015; 7(12): 6707-15.
[http://dx.doi.org/10.3390/v7122967] [PMID: 26694453] -43Ramakrishnaiah V, Thumann C, Fofana I, et al. Exosome-mediated transmission of hepatitis C virus between human hepatoma Huh7.5 cells. Proc Natl Acad Sci USA 2013; 110(32): 13109-13.
[http://dx.doi.org/10.1073/pnas.1221899110] [PMID: 23878230] ]. The mobility of vesicles in the endosomal pathway as they maneuver through the plasma membrane offers numerous chances for a viral disturbance. Viruses can penetrate the endosomal pathway by entry and viral fusion inside of the cell.
Current studies have shown that hepatocyte-derived exosomes that contain HCV RNA can stimulate innate immune cells [43Ramakrishnaiah V, Thumann C, Fofana I, et al. Exosome-mediated transmission of hepatitis C virus between human hepatoma Huh7.5 cells. Proc Natl Acad Sci USA 2013; 110(32): 13109-13.
[http://dx.doi.org/10.1073/pnas.1221899110] [PMID: 23878230] ]. HCV can infect specific target cells in the form of cell-free viruses and through cellular contact [47Liu Z, Zhang X, Yu Q, He JJ. Exosome-associated hepatitis C virus in cell cultures and patient plasma. Biochem Biophys Res Commun 2014; 455(3-4): 218-22.
[http://dx.doi.org/10.1016/j.bbrc.2014.10.146] [PMID: 25449270] ]. In 2013, Ramakrishnaiah et al. reported that HCV infection can be spread by exosomes between hepatocyte-like cells and can create a productive infection [43Ramakrishnaiah V, Thumann C, Fofana I, et al. Exosome-mediated transmission of hepatitis C virus between human hepatoma Huh7.5 cells. Proc Natl Acad Sci USA 2013; 110(32): 13109-13.
[http://dx.doi.org/10.1073/pnas.1221899110] [PMID: 23878230] ]. Another study was reported by Dreux and colleagues [48Dreux M, Garaigorta U, Boyd B, et al. Short-range exosomal transfer of viral RNA from infected cells to plasmacytoid dendritic cells triggers innate immunity. Cell Host Microbe 2012; 12(4): 558-70.
[http://dx.doi.org/10.1016/j.chom.2012.08.010] [PMID: 23084922] ]. They demonstrated that hepatocyte-derived exosomes containing viral RNA can prompt IFN-α production in plasmacytoid dendritic cells [48Dreux M, Garaigorta U, Boyd B, et al. Short-range exosomal transfer of viral RNA from infected cells to plasmacytoid dendritic cells triggers innate immunity. Cell Host Microbe 2012; 12(4): 558-70.
[http://dx.doi.org/10.1016/j.chom.2012.08.010] [PMID: 23084922] ]. In 2014, Liu et al. reported the presence of HCV in exosome-free and exosome-related forms [47Liu Z, Zhang X, Yu Q, He JJ. Exosome-associated hepatitis C virus in cell cultures and patient plasma. Biochem Biophys Res Commun 2014; 455(3-4): 218-22.
[http://dx.doi.org/10.1016/j.bbrc.2014.10.146] [PMID: 25449270] ]. The group revealed that exosome-associated HCV was infectious and resistant to neutralization by an anti-HCV neutralizing antibody [47Liu Z, Zhang X, Yu Q, He JJ. Exosome-associated hepatitis C virus in cell cultures and patient plasma. Biochem Biophys Res Commun 2014; 455(3-4): 218-22.
[http://dx.doi.org/10.1016/j.bbrc.2014.10.146] [PMID: 25449270] ]. In addition, they revealed that more exosome-associated HCV was present compared to exosome-free HCV identified in the plasma of HCV-infected patients [47Liu Z, Zhang X, Yu Q, He JJ. Exosome-associated hepatitis C virus in cell cultures and patient plasma. Biochem Biophys Res Commun 2014; 455(3-4): 218-22.
[http://dx.doi.org/10.1016/j.bbrc.2014.10.146] [PMID: 25449270] ]. Therefore, this evidence suggests that exosome-associated HCV serves as an alternate form for HCV transmission and infection [47Liu Z, Zhang X, Yu Q, He JJ. Exosome-associated hepatitis C virus in cell cultures and patient plasma. Biochem Biophys Res Commun 2014; 455(3-4): 218-22.
[http://dx.doi.org/10.1016/j.bbrc.2014.10.146] [PMID: 25449270] ].
3. EXOSOMES AND PROTEIN TRAFFICKING
Several viruses express membrane proteins [49Lodish H, Berk A, Zipursky SL, et al. Viruses: structure, function, and uses. Molecular cell biology. (4th ed). New York, NY: W. H. Freeman 2000.]. Membrane proteins in enveloped viruses are often structural constituents of the virus that facilitate the important task of membrane fusion and receptor recognition. The activities of these proteins entail that they are copied properly in infected cells. Often, these cataloging events rely on the ability of the virus to make a network with the cellular trafficking machinery and copy cellular protein trafficking signals. Notably, modification or loss of these signals can impact viral pathogenesis and infectivity [50Byland R, Marsh M. Trafficking of viral membrane proteins. Curr Top Microbiol Immunol 2005; 285: 219-54.
[http://dx.doi.org/10.1007/3-540-26764-6_8] [PMID: 15609506] ].
Exosomes found in body fluids play an essential role in exchanging information among cells. There are three mechanisms of contact between exosomes and their recipient cells. First, the transmembrane proteins of exosomes make a network directly with the signaling receptors of target cells [51Munich S, Sobo-Vujanovic A, Buchser WJ, Beer-Stolz D, Vujanovic NL. Dendritic cell exosomes directly kill tumor cells and activate natural killer cells via TNF superfamily ligands. OncoImmunology 2012; 1(7): 1074-83.
[http://dx.doi.org/10.4161/onci.20897] [PMID: 23170255] ]. Secondly, the exosomes join with the plasma membrane of recipient cells and transport their content inside of the cytosol [52Mulcahy LA, Pink RC, Carter DR. Routes and mechanisms of extracellular vesicle uptake. J Extracell Vesicles 2014; 3: 3.
[http://dx.doi.org/10.3402/jev.v3.24641] [PMID: 25143819] ]. Third, the exosomes are incorporated into the recipient cells where they have two possible fates. In the first possible fate, engulfed exosomes may join inside of the endosomes and go through transcytosis. This process allows the exosomes to move across the recipient cells and be released into neighboring cells. If the second possible fate occurs, endosomes can fuse with engulfed exosomes and undergo degradation within the lysosomes [52Mulcahy LA, Pink RC, Carter DR. Routes and mechanisms of extracellular vesicle uptake. J Extracell Vesicles 2014; 3: 3.
[http://dx.doi.org/10.3402/jev.v3.24641] [PMID: 25143819] , 53Tian T, Zhu YL, Hu FH, Wang YY, Huang NP, Xiao ZD. Dynamics of exosome internalization and trafficking. J Cell Physiol 2013; 228(7): 1487-95.
[http://dx.doi.org/10.1002/jcp.24304] [PMID: 23254476] ]. Studies have recently shown the factors that have influenced the internalization of exosomes in recipient cells. Koumangoye et al. detected that disruption of exosomal lipid rafts leads to the inhibition of internalization of exosomes [27Zhang J, Li S, Li L, et al. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genomics Proteomics Bioinformatics 2015; 13(1): 17-24.
[http://dx.doi.org/10.1016/j.gpb.2015.02.001] [PMID: 25724326] , 54Koumangoye RB, Sakwe AM, Goodwin JS, Patel T, Ochieng J. Detachment of breast tumor cells induces rapid secretion of exosomes which subsequently mediate cellular adhesion and spreading. PLoS One 2011; 6(9): e24234.
[http://dx.doi.org/10.1371/journal.pone.0024234] [PMID: 21915303] ]. The group also showed that annexins were necessary for the uptake of exosomes in the breast carcinoma cell line BT-549 [27Zhang J, Li S, Li L, et al. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genomics Proteomics Bioinformatics 2015; 13(1): 17-24.
[http://dx.doi.org/10.1016/j.gpb.2015.02.001] [PMID: 25724326] , 54Koumangoye RB, Sakwe AM, Goodwin JS, Patel T, Ochieng J. Detachment of breast tumor cells induces rapid secretion of exosomes which subsequently mediate cellular adhesion and spreading. PLoS One 2011; 6(9): e24234.
[http://dx.doi.org/10.1371/journal.pone.0024234] [PMID: 21915303] ]. Escrevente et al. described the importance of protease K and exosome-mediated uptake of exosomes derived from the human ovarian carcinoma cell line SKOV3 [27Zhang J, Li S, Li L, et al. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genomics Proteomics Bioinformatics 2015; 13(1): 17-24.
[http://dx.doi.org/10.1016/j.gpb.2015.02.001] [PMID: 25724326] , 55Escrevente C, Keller S, Altevogt P, Costa J. Interaction and uptake of exosomes by ovarian cancer cells. BMC Cancer 2011; 11: 108.
[http://dx.doi.org/10.1186/1471-2407-11-108] [PMID: 21439085] ].
Exosome trafficking allows tetraspanins and a variety of cellular constituents, such as major histocompatibility complex class II (MHC-II) molecules to maneuver through the cell. The process of “back fusion” can aid in the entry of viral proteins that are being generated inside the cell and targeted for the endosomal compartment [44Nour AM, Modis Y. Endosomal vesicles as vehicles for viral genomes. Trends Cell Biol 2014; 24(8): 449-54.
[http://dx.doi.org/10.1016/j.tcb.2014.03.006] [PMID: 24746011] ]. Viruses, such as herpesviruses and other DNA viruses that require proximity to the nucleus and endoplasmic reticulum, will also follow this route and possibly have components incorporated into the exosomes [42Anderson MR, Kashanchi F, Jacobson S. Exosomes in viral disease. Neurotherapeutics 2016; 13(3): 535-46.
[http://dx.doi.org/10.1007/s13311-016-0450-6] [PMID: 27324390] ] (Table 1).
4. EXOSOMES AND VIRAL INFECTIONS
Exosomes have several characteristics that are like some viruses. These characteristics include biogenesis, molecular properties uptake by cells, and exosome-mediated intercellular transfer of functional RNAs, mRNAs, and cellular proteins [12Meckes DG Jr, Raab-Traub N. Microvesicles and viral infection. J Virol 2011; 85(24): 12844-54.
[http://dx.doi.org/10.1128/JVI.05853-11] [PMID: 21976651] ]. The differences between exosomes and some viruses include self-replication after infection of new cells, temporary regulated viral expression, and the complexity of viral entry [56Alenquer M, Amorim MJ. Exosome Biogenesis, Regulation, and Function in Viral Infection. Viruses 2015; 7(9): 5066-83.
[http://dx.doi.org/10.3390/v7092862] [PMID: 26393640] ]. Several virus-infected cells secrete exosomes that vary from their virion counterparts but may consist of numerous RNAs and viral proteins. To some degree, exosomes have been evaluated for their content and attributes during viral infection [12Meckes DG Jr, Raab-Traub N. Microvesicles and viral infection. J Virol 2011; 85(24): 12844-54.
[http://dx.doi.org/10.1128/JVI.05853-11] [PMID: 21976651] ]. However, more investigation is needed, as it relates to DNA viruses. Table 1 summarizes the relationship of DNA and RNA virus infection and exosomes.
5. RETROVIRUSES
Retroviruses are enveloped RNA viruses that bud at the plasma membrane of infected cells [57Pelchen-Matthews A, Raposo G, Marsh M. Endosomes, exosomes and Trojan viruses. Trends Microbiol 2004; 12(7): 310-6.
[http://dx.doi.org/10.1016/j.tim.2004.05.004] [PMID: 15223058] ]. They utilize reverse transcriptase to change their single-stranded RNA into double-stranded DNA. This allows the reverse transcriptase to become integrated inside the genome of the cells which have been infected [58Hu WS, Hughes SH. HIV-1 reverse transcription. Cold Spring Harb Perspect Med 2012; 2(10): a006882.
[http://dx.doi.org/10.1101/cshperspect.a006882] [PMID: 23028129] ]. All retroviruses are comprised of three key coding domains i) gag, ii) pol, and iii) env [59Coffin JM, Hughes SH, Varmus HE. The interactions of retroviruses and their hosts.Retroviruses 1997.]. Gag guides the synthesis of internal virion proteins that make up the nucleoprotein structures, the matrix, and the capsid. Pol comprises the information that is needed for integrase and reverse transcriptase and enzymes. Env descends the transmembrane and surface constituents of the viral envelope protein. Pro is a smaller coding domain. It encodes the virion protease in all retroviruses. In most cases, simple retroviruses only carry this primary information. However, complex retroviruses code for non-virion proteins that develop from various spliced messages [59Coffin JM, Hughes SH, Varmus HE. The interactions of retroviruses and their hosts.Retroviruses 1997.].
Retroviruses are “retro” because they reverse the route of the standard gene copying procedures [60Anson DS. The use of retroviral vectors for gene therapy-what are the risks? A review of retroviral pathogenesis and its relevance to retroviral vector-mediated gene delivery. Genet Vaccines Ther 2004; 2(1): 9.
[http://dx.doi.org/10.1186/1479-0556-2-9] [PMID: 15310406] ]. According to the Trojan exosome hypothesis, retroviruses could be exosomes that developed upon the mutation of a structural gag gene [61Izquierdo-Useros N, Puertas MC, Borràs FE, Blanco J, Martinez-Picado J. Exosomes and retroviruses: The chicken or the egg? Cell Microbiol 2011; 13(1): 10-7.
[http://dx.doi.org/10.1111/j.1462-5822.2010.01542.x] [PMID: 21054740] ]. The gag gene is encoded by an integrated retrotransposon that directs viral biosynthesis to the exosome pathway [61Izquierdo-Useros N, Puertas MC, Borràs FE, Blanco J, Martinez-Picado J. Exosomes and retroviruses: The chicken or the egg? Cell Microbiol 2011; 13(1): 10-7.
[http://dx.doi.org/10.1111/j.1462-5822.2010.01542.x] [PMID: 21054740] ]. The most studied retrovirus that infects humans is HIV-1 [62Sharp PM, Hahn BH. Origins of HIV and the AIDS pandemic. Cold Spring Harb Perspect Med 2011; 1(1): a006841.
[http://dx.doi.org/10.1101/cshperspect.a006841] [PMID: 22229120] ].
6. HUMAN IMMUNODEFICIENCY VIRUS
The potential functions of exosomes secreted during HIV-1 and HIV-2 infections are just beginning to be explored. HIV-1 and HIV-2 share the same modes of transmission [63Blut A. Human Immunodeficiency Virus (HIV). Transfus Med Hemother 2016; 43(3): 203-22.
[http://dx.doi.org/10.1159/000445852] [PMID: 27403093] ]. Immunodeficiency develops gradually in people that are infected with HIV-2. HIV-2 infected people are less infectious in the early phase of infection as compared to those infected with HIV-1. As the disease progresses, HIV-2 increases. However, in the case of HIV-1, as the disease progresses, HIV-1 decreases [63Blut A. Human Immunodeficiency Virus (HIV). Transfus Med Hemother 2016; 43(3): 203-22.
[http://dx.doi.org/10.1159/000445852] [PMID: 27403093] , 64Cohen MS, Hellmann N, Levy JA, DeCock K, Lange J. The spread, treatment, and prevention of HIV-1: Evolution of a global pandemic. J Clin Invest 2008; 118(4): 1244-54.
[http://dx.doi.org/10.1172/JCI34706] [PMID: 18382737] ].
HIV-1 and HIV-2 are two distinct viruses that infect monocytes/macrophages and T lymphocytes. These types of cells express CD4, a cell surface type I transmembrane glycoprotein, which is the major receptor for HIV [65de Carvalho JV, de Castro RO, da Silva EZ, et al. Nef neutralizes the ability of exosomes from CD4+ T cells to act as decoys during HIV-1 infection. PLoS One 2014; 9(11): e113691.
[http://dx.doi.org/10.1371/journal.pone.0113691] [PMID: 25423108] ]. Host cell infection starts with the involvement of the gp120 subunit of the viral envelope (Env) glycoprotein-network through CD4. This involvement leads to variations in gp120 that increase its affinity to chemokine co-receptors, CXCR4 or CCR5. Co-receptor binding to Env initiates changes in the gp41 subunit. These changes stimulate fusion and tethering of the Env to the cell plasma membrane [66Ray N, Doms RW. HIV-1 coreceptors and their inhibitors. Curr Top Microbiol Immunol 2006; 303: 97-120.
[http://dx.doi.org/10.1007/978-3-540-33397-5_5] [PMID: 16570858] , 67Melikyan GB. HIV entry: A game of hide-and-fuse? Curr Opin Virol 2014; 4: 1-7.
[http://dx.doi.org/10.1016/j.coviro.2013.09.004] [PMID: 24525288] ]. CD4-Env collaboration is essential for viral entry. Therefore, the use of participant particles that target this interaction [65de Carvalho JV, de Castro RO, da Silva EZ, et al. Nef neutralizes the ability of exosomes from CD4+ T cells to act as decoys during HIV-1 infection. PLoS One 2014; 9(11): e113691.
[http://dx.doi.org/10.1371/journal.pone.0113691] [PMID: 25423108] ] or the downregulation of CD4 surface expression [68Benson RE, Sanfridson A, Ottinger JS, Doyle C, Cullen BR. Downregulation of cell-surface CD4 expression by simian immunodeficiency virus Nef prevents viral super infection. J Exp Med 1993; 177(6): 1561-6.
[http://dx.doi.org/10.1084/jem.177.6.1561] [PMID: 8098729] ] can inhibit HIV-1 infection.
Exosomes display similar molecular and structural characteristics with HIV-1 and HIV-2. They are both enclosed by a lipid bilayer. Their size and density [69Kowal J, Tkach M, Théry C. Biogenesis and secretion of exosomes. Curr Opin Cell Biol 2014; 29: 116-25.
[http://dx.doi.org/10.1016/j.ceb.2014.05.004] [PMID: 24959705] ] vary from 50 to 150 nm in diameter [70Conde-Vancells J, Rodriguez-Suarez E, Embade N, et al. Characterization and comprehensive proteome profiling of exosomes secreted by hepatocytes. J Proteome Res 2008; 7(12): 5157-66.
[http://dx.doi.org/10.1021/pr8004887] [PMID: 19367702] ] and 1.13 to 1.21 g/mL [71Théry C, Boussac M, Véron P, et al. Proteomic analysis of dendritic cell-derived exosomes: A secreted subcellular compartment distinct from apoptotic vesicles. J Immunol 2001; 166(12): 7309-18.
[http://dx.doi.org/10.4049/jimmunol.166.12.7309] [PMID: 11390481] ], correspondingly. Also, they are both composed of RNA species [61Izquierdo-Useros N, Puertas MC, Borràs FE, Blanco J, Martinez-Picado J. Exosomes and retroviruses: The chicken or the egg? Cell Microbiol 2011; 13(1): 10-7.
[http://dx.doi.org/10.1111/j.1462-5822.2010.01542.x] [PMID: 21054740] ], carbohydrates [72Krishnamoorthy L, Bess JW Jr, Preston AB, Nagashima K, Mahal LK. HIV-1 and microvesicles from T cells share a common glycome, arguing for a common origin. Nat Chem Biol 2009; 5(4): 244-50.
[http://dx.doi.org/10.1038/nchembio.151] [PMID: 19234452] ], lipids [73Wubbolts R, Leckie RS, Veenhuizen PT, et al. Proteomic and biochemical analyses of human B cell-derived exosomes. Potential implications for their function and multivesicular body formation. J Biol Chem 2003; 278(13): 10963-72.
[http://dx.doi.org/10.1074/jbc.M207550200] [PMID: 12519789] ], and proteins [14Théry C, Zitvogel L, Amigorena S. Exosomes: Composition, biogenesis and function. Nat Rev Immunol 2002; 2(8): 569-79.
[http://dx.doi.org/10.1038/nri855] [PMID: 12154376] , 74Ott DE. Cellular proteins detected in HIV-1. Rev Med Virol 2008; 18(3): 159-75.
[http://dx.doi.org/10.1002/rmv.570] [PMID: 18265424] ]. Exosomes derived from HIV-infected cells are enhanced with viral RNAs [75Valadi H, Ekström K, Bossios A, Sjöstrand M, Lee JJ, Lötvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol 2007; 9(6): 654-9.
[http://dx.doi.org/10.1038/ncb1596] [PMID: 17486113] , 76Pegtel DM, Cosmopoulos K, Thorley-Lawson DA, et al. Functional delivery of viral miRNAs via exosomes. Proc Natl Acad Sci USA 2010; 107(14): 6328-33.
[http://dx.doi.org/10.1073/pnas.0914843107] [PMID: 20304794] ] and Nef protein [77Lenassi M, Cagney G, Liao M, et al. HIV Nef is secreted in exosomes and triggers apoptosis in bystander CD4+ T cells. Traffic 2010; 11(1): 110-22.
[http://dx.doi.org/10.1111/j.1600-0854.2009.01006.x] [PMID: 19912576] ].
Because of these comparisons, studies have shown that HIV-1 can be produced by the identical pathways of exosome biogenesis [39Madison MN, Jones PH, Okeoma CM. Exosomes in human semen restrict HIV-1 transmission by vaginal cells and block intravaginal replication of LP-BM5 murine AIDS virus complex. Virology 2015; 482: 189-201.
[http://dx.doi.org/10.1016/j.virol.2015.03.040] [PMID: 25880110] , 61Izquierdo-Useros N, Puertas MC, Borràs FE, Blanco J, Martinez-Picado J. Exosomes and retroviruses: The chicken or the egg? Cell Microbiol 2011; 13(1): 10-7.
[http://dx.doi.org/10.1111/j.1462-5822.2010.01542.x] [PMID: 21054740] ]. In favor of the Trojan exosome hypothesis, HIV-1 recruits constituents of the host ESCRT machinery to the appropriate sight of viral budding [12Meckes DG Jr, Raab-Traub N. Microvesicles and viral infection. J Virol 2011; 85(24): 12844-54.
[http://dx.doi.org/10.1128/JVI.05853-11] [PMID: 21976651] , 39Madison MN, Jones PH, Okeoma CM. Exosomes in human semen restrict HIV-1 transmission by vaginal cells and block intravaginal replication of LP-BM5 murine AIDS virus complex. Virology 2015; 482: 189-201.
[http://dx.doi.org/10.1016/j.virol.2015.03.040] [PMID: 25880110] ]. The collaboration between tetraspanin and HIV-1 Gag protein proposes that HIV-1 may use lipid raft micro-domains rich in tetraspanins for virus assembly [78Grigorov B, Attuil-Audenis V, Perugi F, et al. A role for CD81 on the late steps of HIV-1 replication in a chronically infected T cell line. Retrovirology 2009; 6: 28.
[http://dx.doi.org/10.1186/1742-4690-6-28] [PMID: 19284574] ]. Tetraspanins CD63 and CD81 are located on the surface of some exosomes. CD63 and CD81 are known to be involved with HIV-1 infectivity [79Raymond AD, Campbell-Sims TC, Khan M, et al. HIV Type 1 Nef is released from infected cells in CD45(+) microvesicles and is present in the plasma of HIV-infected individuals. AIDS Res Hum Retroviruses 2011; 27(2): 167-78.
[http://dx.doi.org/10.1089/aid.2009.0170] [PMID: 20964480] ]. In addition, exosomes and HIV-1 express sialyllactose-containing gangliosides. Studies have shown that sialic-acid-binding immunoglobulin-like lectins (Siglecs)-1 interact with sialyllactose-containing ganglioside on exosomes and HIV-1. Siglecs-1 stimulates mature dendritic cell (mDC) capture and storage of both exosomes and HIV-1 in mDCs. This facilitates trans infection of T cells by mDCs [80Izquierdo-Useros N, Lorizate M, Puertas MC, et al. Siglec-1 is a novel dendritic cell receptor that mediates HIV-1 trans-infection through recognition of viral membrane gangliosides. PLoS Biol 2012; 10(12): e1001448.
[http://dx.doi.org/10.1371/journal.pbio.1001448] [PMID: 23271952] ].
Studies have shown that exosomes that are released from infected cells contain co-receptors for HIV-1 which can improve virus entry inside cells [81Madison MN, Okeoma CM. Exosomes: Implications in HIV-1 Pathogenesis. Viruses 2015; 7(7): 4093-118.
[http://dx.doi.org/10.3390/v7072810] [PMID: 26205405] ]. Expression of the viral Nef protein modifies the endosomal network by maximizing the amount of MVBs, endosomes, and lysosomes. Nef is a protein that is coded by HIV-1 and HIV-2 genomes. The Nef protein modulates protein trafficking and signal transduction mechanisms in infected cells (39). Recent studies have shown that Nef is displayed in exosomes that are derived from cells. Studies have also shown that Nef-containing exosomes prompted apoptosis in CD4+ T cells. Thus, exosomal Nef can participate in HIV-1 and HIV-2 pathogenesis by aiding in the reduction of CD4+ T cells [12Meckes DG Jr, Raab-Traub N. Microvesicles and viral infection. J Virol 2011; 85(24): 12844-54.
[http://dx.doi.org/10.1128/JVI.05853-11] [PMID: 21976651] ].
Nef promotes MHC-I molecules and the downregulation of CD4’s cell surface. Nef attaches itself to the cytosolic tail of MHC-I and CD4+. Due to this attachment, it disturbs the intracellular trafficking of these proteins through reformed mechanisms [82Roeth JF, Williams M, Kasper MR, Filzen TM, Collins KL. HIV-1 Nef disrupts MHC-I trafficking by recruiting AP-1 to the MHC-I cytoplasmic tail. J Cell Biol 2004; 167(5): 903-13.
[http://dx.doi.org/10.1083/jcb.200407031] [PMID: 15569716] ]. Nef stimulates CD4 endocytosis via clathrin-coated vesicles. This allows the mechanism to form a tripartite complex in the clathrin pits with the adaptor protein (AP) complex 2 and CD4 [83Chaudhuri R, Mattera R, Lindwasser OW, Robinson MS, Bonifacino JS. A basic patch on alpha-adaptin is required for binding of human immunodeficiency virus type 1 Nef and cooperative assembly of a CD4-Nef-AP-2 complex. J Virol 2009; 83(6): 2518-30.
[http://dx.doi.org/10.1128/JVI.02227-08] [PMID: 19129443] ] at the plasma membrane. In contrast , Nef inhibits MHC-I from attaining the plasma membrane by stimulating the retention of these molecules inside the Golgi apparatus by interacting with AP-1 [82Roeth JF, Williams M, Kasper MR, Filzen TM, Collins KL. HIV-1 Nef disrupts MHC-I trafficking by recruiting AP-1 to the MHC-I cytoplasmic tail. J Cell Biol 2004; 167(5): 903-13.
[http://dx.doi.org/10.1083/jcb.200407031] [PMID: 15569716] ]. Also, Nef can prompt MHC-I internalization by initiating a signal transduction pathway that involves the assembly of a kinase cascade [84Hung CH, Thomas L, Ruby CE, et al. HIV-1 Nef assembles a Src family kinase-ZAP-70/Syk-PI3K cascade to downregulate cell-surface MHC-I. Cell Host Microbe 2007; 1(2): 121-33.
[http://dx.doi.org/10.1016/j.chom.2007.03.004] [PMID: 18005690] ]. Both replicas believe that Nef transmits MHC-I molecules toward MVBs and late endosomes inside the endolysosomal pathway. In addition, MHC-I and CD4 are directed to MVBs upon Nef expression [85Lubben NB, Sahlender DA, Motley AM, Lehner PJ, Benaroch P, Robinson MS. HIV-1 Nef-induced down-regulation of MHC class I requires AP-1 and clathrin but not PACS-1 and is impeded by AP-2. Mol Biol Cell 2007; 18(9): 3351-65.
[http://dx.doi.org/10.1091/mbc.e07-03-0218] [PMID: 17581864] ].
7. EXOSOMES AUGMENT HIV-1 ENTRY
As previously mentioned, exosomes can play a vital role in the entry of HIV-1 viral infection. HIV-1 utilizes primary and secondary cellular receptors in order to invade host cells [86Sims B, Farrow AL, Williams SD, et al. Role of TIM-4 in exosome-dependent entry of HIV-1 into human immune cells. Int J Nanome 2017; 12: 4823-33.
[http://dx.doi.org/10.2147/IJN.S132762] [PMID: 28740388] ]. These receptors facilitate precise, high-affinity collaborations with viral entry proteins. In addition, these receptors prime the entry protein for consequent phases in the viral entry method [87Jemielity S, Wang JJ, Chan YK, et al. TIM-family proteins promote infection of multiple enveloped viruses through virion-associated phosphatidylserine. PLoS Pathog 2013; 9(3): e1003232.
[http://dx.doi.org/10.1371/journal.ppat.1003232] [PMID: 23555248] ]. HIV-1 has an Env that it secures when it leaves the cells. In order to infect cells, the HIV-1 Env protein fuses near the primary cellular receptor and then to the cellular co-receptor. Sequentially, this binding causes fusion between the host and viral cell membranes which aid in the initiation of infection [88Wilen CB, Tilton JC, Doms RW. HIV: Cell binding and entry. Cold Spring Harb Perspect Med 2012; 2(8): a006866.
[http://dx.doi.org/10.1101/cshperspect.a006866] [PMID: 22908191] ].
HIV-1 uses human T-cell immunoglobin mucin (TIM) proteins to augment viral entry. TIMs are a group of proteins (TIM-1, TIM-3, and TIM-4) that promote phagocytosis of apoptotic cells [87Jemielity S, Wang JJ, Chan YK, et al. TIM-family proteins promote infection of multiple enveloped viruses through virion-associated phosphatidylserine. PLoS Pathog 2013; 9(3): e1003232.
[http://dx.doi.org/10.1371/journal.ppat.1003232] [PMID: 23555248] ]. The interaction of dying cells and enveloped viruses is mediated via phosphatidylserine (PtdSer). PtdSer is a phospholipid that acts as an assistant constituent for viral fusion, using [86Sims B, Farrow AL, Williams SD, et al. Role of TIM-4 in exosome-dependent entry of HIV-1 into human immune cells. Int J Nanome 2017; 12: 4823-33.
[http://dx.doi.org/10.2147/IJN.S132762] [PMID: 28740388] , 89Arenaccio C, Chiozzini C, Columba-Cabezas S, et al. Exosomes from human immunodeficiency virus type 1 (HIV-1)-infected cells license quiescent CD4+ T lymphocytes to replicate HIV-1 through a Nef- and ADAM17-dependent mechanism. J Virol 2014; 88(19): 11529-39.
[http://dx.doi.org/10.1128/JVI.01712-14] [PMID: 25056899] ] residues that are exposed to the cellular and viral membranes [87Jemielity S, Wang JJ, Chan YK, et al. TIM-family proteins promote infection of multiple enveloped viruses through virion-associated phosphatidylserine. PLoS Pathog 2013; 9(3): e1003232.
[http://dx.doi.org/10.1371/journal.ppat.1003232] [PMID: 23555248] ]. In previous reports, Sims et al., showed that neural stem cell (NSC)-derived exosomes have TIM-4 protein. A few years later, the group showed that TIM-4 is involved in HIV-1 exosome-dependent cellular entry mechanism. Also, they showed that HIV-1, which consists of large quantities of PtdSer, could bind to PtdSer receptors on exosomes, such as TIM-4. Thus, they concluded that exosomes from multiple sources, such as breast milk, blood, NSC, and human epithelial lung cells (A549), increased HIV-1 cellular entry. Sims and the group also demonstrated that exosome-mediated entry was effectively blocked using a TIM-4 antibody [86Sims B, Farrow AL, Williams SD, et al. Role of TIM-4 in exosome-dependent entry of HIV-1 into human immune cells. Int J Nanome 2017; 12: 4823-33.
[http://dx.doi.org/10.2147/IJN.S132762] [PMID: 28740388] ].
Fig. (1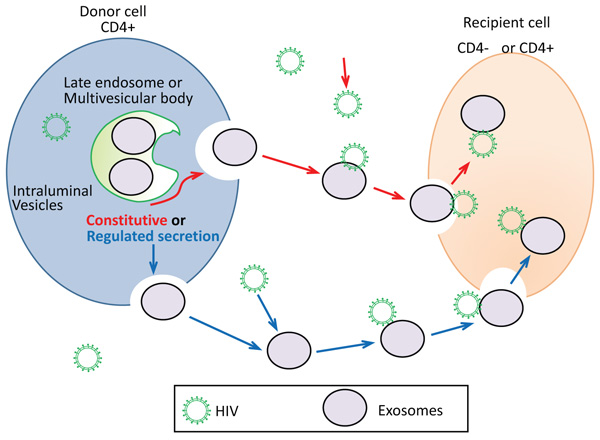 ) depicts the proposed mechanism of putative exosome-mediate HIV-1 entry in CD+4 or CD-4 recipient cells. As previously mentioned, the formation of exosomes occurs when the late endocytotic constituents join with the plasma membrane. This process can be either constitutive or regulated (Fig. 1). Constitutive release of exosomes represent secretion of exosomes that is constant. Whereas, regulated exosome release represents exosome release that is the result of a stimulus or signal. Constitutive secretion of exosomes is seen in most types of cells, such as epithelial cells, EBV-transformed B cells, and immature dendritic cells [4van Niel G, Raposo G, Candalh C, et al. Intestinal epithelial cells secrete exosome-like vesicles. Gastroenterology 2001; 121(2): 337-49.
) depicts the proposed mechanism of putative exosome-mediate HIV-1 entry in CD+4 or CD-4 recipient cells. As previously mentioned, the formation of exosomes occurs when the late endocytotic constituents join with the plasma membrane. This process can be either constitutive or regulated (Fig. 1). Constitutive release of exosomes represent secretion of exosomes that is constant. Whereas, regulated exosome release represents exosome release that is the result of a stimulus or signal. Constitutive secretion of exosomes is seen in most types of cells, such as epithelial cells, EBV-transformed B cells, and immature dendritic cells [4van Niel G, Raposo G, Candalh C, et al. Intestinal epithelial cells secrete exosome-like vesicles. Gastroenterology 2001; 121(2): 337-49.
[http://dx.doi.org/10.1053/gast.2001.26263] [PMID: 11487543] , 90Raposo G, Nijman HW, Stoorvogel W, et al. B lymphocytes secrete antigen-presenting vesicles. J Exp Med 1996; 183(3): 1161-72.
[http://dx.doi.org/10.1084/jem.183.3.1161] [PMID: 8642258] , 91Théry C, Regnault A, Garin J, et al. Molecular characterization of dendritic cell-derived exosomes. Selective accumulation of the heat shock protein hsc73. J Cell Biol 1999; 147(3): 599-610.
[http://dx.doi.org/10.1083/jcb.147.3.599] [PMID: 10545503] ]. Other cells can have regulated secretion of exosomes such as mast cells and T cells. Calcium has been shown to be a key regulator of this process [92Raposo G, Tenza D, Mecheri S, Peronet R, Bonnerot C, Desaymard C. Accumulation of major histocompatibility complex class II molecules in mast cell secretory granules and their release upon degranulation. Mol Biol Cell 1997; 8(12): 2631-45.
[http://dx.doi.org/10.1091/mbc.8.12.2631] [PMID: 9398681] , 93Blanchard N, Lankar D, Faure F, et al. TCR activation of human T cells induces the production of exosomes bearing the TCR/CD3/zeta complex. J Immunol 2002; 168(7): 3235-41.
[http://dx.doi.org/10.4049/jimmunol.168.7.3235] [PMID: 11907077] ]. Activated CD4+ T cells were initially described to secrete EVs which were associated with FasL and APO2L (68). The secretion occurred in a putative mechanism to maintain immune tolerance and T cell homeostasis through the apoptosis of targeted cells [94Martínez-Lorenzo MJ, Anel A, Gamen S, et al. Activated human T cells release bioactive Fas ligand and APO2 ligand in microvesicles. J Immunol 1999; 163(3): 1274-81.
[PMID: 10415024] ]. HIV-1 infects CD4+ T cells and evades the host immune response. It also stimulates an inflammatory immune response which can cause cell exhaustion [95Soares H. HIV-1 Intersection with CD4 T Cell Vesicle Exocytosis: Intercellular Communication Goes Viral. Front Immunol 2014; 5: 454.
[http://dx.doi.org/10.3389/fimmu.2014.00454] [PMID: 25295039] ]. It has been well noted that HIV-1 enters T cells by binding and engagement of co-receptors and CD4. However, based on the findings of Sims et al. [86Sims B, Farrow AL, Williams SD, et al. Role of TIM-4 in exosome-dependent entry of HIV-1 into human immune cells. Int J Nanome 2017; 12: 4823-33.
[http://dx.doi.org/10.2147/IJN.S132762] [PMID: 28740388] ], it is speculated that HIV-1 can interact and bind secreted exosomes and enter into CD4+ and CD4- negative cells (Fig. 1). Sims et. al proposed the mechanism of interaction of exosomes and HIV to be mediated by TIM-4, CD9, and/or CD81. It is possible that this interaction between exosomes and HIV-1 is specific to the origin of the exosome; therefore, further studies are required [96Sims B, Farrow AL, Williams SD, Bansal A, Krendelchtchikov A, Matthews QL. Tetraspanin blockage reduces exosome-mediated HIV-1 entry. Arch Virol 2018; 163(6): 1683-3.
[http://dx.doi.org/10.1007/s00705-018-3737-6] [PMID: 29429034] , 97Gu L, Sims B, Krendelchtchikov A, Tabengwa E, Matthews QL. Differential binding of the HIV-1 envelope to phosphatidylserine receptors. Biochim Biophys Acta 2017; 1859(10): 1962-6.
[http://dx.doi.org/10.1016/j.bbamem.2017.06.007] [PMID: 28622976] ].

8. DNA VIRUSES HIJACKING
Viruses are categorized according to the proteins that are encoded in the genome or viral genetic material, such as DNA [49Lodish H, Berk A, Zipursky SL, et al. Viruses: structure, function, and uses. Molecular cell biology. (4th ed). New York, NY: W. H. Freeman 2000.]. DNA viruses are intracellular parasites that can only reproduce in cells. They can program the cell to reproduce the virus using the genes that are obtained inside the viral DNA genome. DNA viruses replicate their genomes with DNA polymerases and RNA polymerases [98Choi KH. Viral polymerases. Adv Exp Med Biol 2012; 726: 267-304.
[http://dx.doi.org/10.1007/978-1-4614-0980-9_12] [PMID: 22297518] ]. With respect to the viral replication cycle, many DNA viruses control gene expression in a “time-ordered” approach. The virus expresses “early genes ” and “late genes.” The early genes interact with the cell to initiate the cell's DNA machinery. The late genes are mostly virion proteins that are essential for virion assembly. Except for poxviruses, all DNA viruses reproduce inside the nucleus of the infected cells. These viruses are dependent on a lesser or greater amount of the cell's DNA machinery. Most cells in the host are in a latent state, in which DNA synthesis is not accessible for viral DNA replication [99Knipe DM. The role of viral and cellular nuclear proteins in herpes simplex virus replication. Adv Virus Res 1989; 37: 85-123.
[http://dx.doi.org/10.1016/S0065-3527(08)60833-7] [PMID: 2557760] ]. The extracellular form of a virus is called a virion. The virion is composed of a group of DNA genes that are sheltered by a protein containing coat termed as capsid. Symmetry and regularity categorize the capsid’s structure. It can invade and bind to cells. In some DNA viruses, the capsid is covered by a membrane, which is made from cellular membranes [49Lodish H, Berk A, Zipursky SL, et al. Viruses: structure, function, and uses. Molecular cell biology. (4th ed). New York, NY: W. H. Freeman 2000.].
One DNA virus, called adenovirus (Ad), binds the cell surface Coxsackie virus and Adenovirus Receptor (CAR) on host cells, resulting in the entry of the virion into the host cell [36Zhang Y, Bergelson JM. Adenovirus receptors. J Virol 2005; 79(19): 12125-31.
[http://dx.doi.org/10.1128/JVI.79.19.12125-12131.2005] [PMID: 16160140] ]. Entry of Ads inside the host cell is started by the knob domain of the fiber protein that binds to the host receptor. Next, a specified motif in the penton base protein makes a network with αv integrin. This stimulates internalization of the adenovirus via clathrin-coated pits, which can result in the entry of the virion inside the host in an endosome. Following internalization, the endosome acidifies which modifies the topology of viruses and causes capsid constituents to separate. As a result, the virion is released into the cytoplasm. The virus is then delivered to the nuclear pore complex where viral gene expression can occur (National Institutes of Health 2002).
For gene therapy approaches, adenovirus serotype 5 (Ad5) has gained increasing attention due to their ability to infect different types of cells. However, natural Ad tropism doesn’t allow Ad to infect cells that are deficient for CAR. In this regard, Sims et al. investigated the role of NSC-derived exosomes to traffic Ad into a CAR-deficient cell line, mouse B cell line A20 [100Sims B, Gu L, Krendelchtchikov A, Matthews QL. Neural stem cell-derived exosomes mediate viral entry. Int J Nanomed 2014; 9: 4893-7.
[http://dx.doi.org/10.2147/IJN.S70999] [PMID: 25364247] ]. This group showed that NSC-derived exosomes facilitated substantial cellular entry of Ad5 in a receptor-independent manner. They showed that TIM-4 found on primary mouse NSC-derived exosomes play an essential role in the cellular entry of Ads. In addition, the group showed that treatment with TIM-4 antibody significantly blocked the exosome-mediated Ad entry [86Sims B, Farrow AL, Williams SD, et al. Role of TIM-4 in exosome-dependent entry of HIV-1 into human immune cells. Int J Nanome 2017; 12: 4823-33.
[http://dx.doi.org/10.2147/IJN.S132762] [PMID: 28740388] , 100Sims B, Gu L, Krendelchtchikov A, Matthews QL. Neural stem cell-derived exosomes mediate viral entry. Int J Nanomed 2014; 9: 4893-7.
[http://dx.doi.org/10.2147/IJN.S70999] [PMID: 25364247] ].
Sims and colleagues hypothesized that exosome-mediated viral entry not only occurs in vitro but occurs in vivo also (unpublished work). To test their hypothesis, Sims et al. co-incubated NSC-derived exosomes with an Ad5 vector (expressing luciferase after infection) and evaluated the ability of exosomes to mediate viral entry. After co-incubation, the Ad5 and exosome complexes were administered intranasally to mice and compared to Ad5 only and PBS. The data illustrated luciferase activity in the mouse group injected with Ad5 only. In addition, there was an increase in luciferase activity in animals injected with Ad and co-incubated with 10 µg/ml of exosomes (Fig. 2A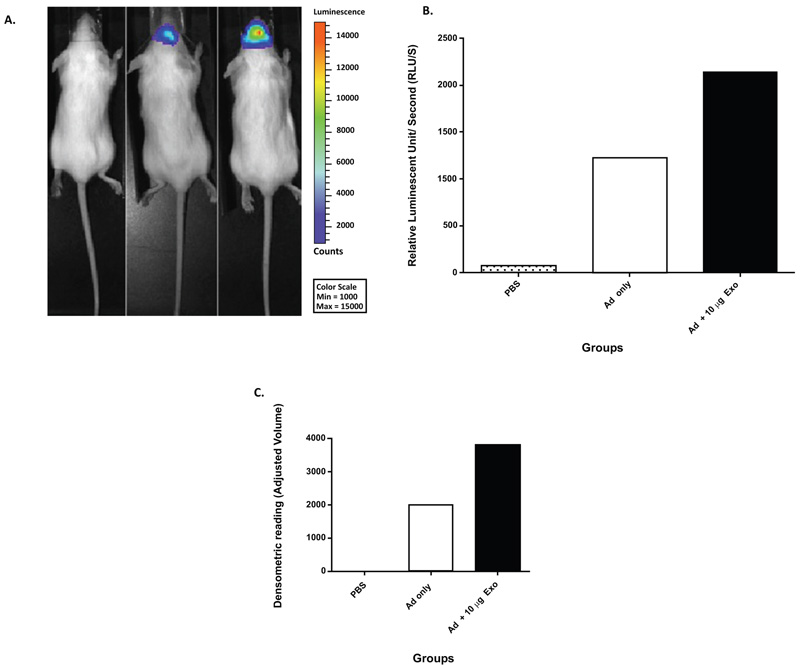 -B). To further confirm these findings, the mouse brains were extracted, sliced, and analyzed by western blot analysis. The results of the western blot analysis were reported via densitometric means (Fig. 2C). This data demonstrated that NSC-derived exosomes could transfer Ad to the brain in an Ad receptor-independent process in vivo. This work could have implications for gene therapy applications targeting the brain.
-B). To further confirm these findings, the mouse brains were extracted, sliced, and analyzed by western blot analysis. The results of the western blot analysis were reported via densitometric means (Fig. 2C). This data demonstrated that NSC-derived exosomes could transfer Ad to the brain in an Ad receptor-independent process in vivo. This work could have implications for gene therapy applications targeting the brain.
 |
Fig. (2) Mouse NSC-derived exosome-mediated in vivo enhancement of Ad infection (unpublished work by Matthews et al.). (A) Mice were injected with PBS, 1x108 Viral Particles (VP) of Ad, or 1x108 VP of Ad co-incubated with 10 µg of exosomes. At 24 hours, post-injection mice were imaged by means of non-invasive luciferase imaging. The imaging results were represented as total luminescent counts. (B) Quantitation of representative mice from Fig. (2A ), data represented as relative luminescent counts per second. (C) Densitometric analysis of western blot analysis of mouse brains at 48 hours post-injection (PBS, 1x108 VP of Ad, or 1x108 VP of Ad co-incubated with 10 µg of exosomes). Anti-adenovirus antibody was used for western blot analysis.
|
Innovative biological research has shown how some viruses hijack cells by copying a signaling indicator to eradicate the body’s resistance. The virus destroys a defensive protein by using cell signals to inhibit it. Viruses replicate themselves by hijacking cellular processes. To replicate, viruses must distribute their DNA inside the nucleus of a cell. This allows a viral infection to cause conflict between the foreign DNA of the virus and the DNA of the host cell.
As a way of penetrating the cell’s defenses, viruses initiate their attack by interacting with cellular proteins. Weitzman et al. showed that the viral protein ICP0 utilizes phosphorylation- a biochemical reaction that is used in the cells to stimulate exchanges among proteins and cell signaling response to DNA damage, in Herpes Simplex Virus-1 (HSV-1) [101Chaurushiya MS, Lilley CE, Aslanian A, et al. Viral E3 ubiquitin ligase-mediated degradation of a cellular E3: Viral mimicry of a cellular phosphorylation mark targets the RNF8 FHA domain. Mol Cell 2012; 46(1): 79-90.
[http://dx.doi.org/10.1016/j.molcel.2012.02.004] [PMID: 22405594] ]. In HSV-1 infection, the phosphorylation signal on ICP0 attracts a cellular DNA damage response protein called RNF8. RNF8 binds to the false signaling marker and is then destroyed. Since RNF8 prevents viral replication, its destruction leaves the cell prone to HSV-1 infection, allowing the virus to take over the cell’s machinery [101Chaurushiya MS, Lilley CE, Aslanian A, et al. Viral E3 ubiquitin ligase-mediated degradation of a cellular E3: Viral mimicry of a cellular phosphorylation mark targets the RNF8 FHA domain. Mol Cell 2012; 46(1): 79-90.
[http://dx.doi.org/10.1016/j.molcel.2012.02.004] [PMID: 22405594] ]. To survive and replicate, the virus binds to the host cell and inserts its RNA or DNA inside the host. The viral DNA takes control of the cell machinery and creates multiple copies of its genomic material thus resulting in several new virions.
9. IMMUNE RESPONSE TO DNA VIRUSES
The immune system defends itself against viral infections by innate and adaptive immune responses [102Nie Y, Wang YY. Innate immune responses to DNA viruses. Protein Cell 2013; 4(1): 1-7.
[http://dx.doi.org/10.1007/s13238-012-2122-6] [PMID: 23307778] ]. The innate immune system serves as the major line of defense against pathogens, which is also needed for preparing for adaptive immunity. Innate immune responses are started by pattern-recognition receptors (PRRs). PRRs identify Pathogen-Associated Molecular Patterns (PAMPs), molecular structures of pathogens. The infection of virus activates a sequence of signaling procedures that lead to proinflammatory cytokines and transcriptional induction of type I interferons (IFNs). Recently, studies have reported vital information pertaining to RNA virus-triggered signaling pathways and the mechanisms of viral RNA recognition. Conversely, it remains uncertain on how DNA virus infection is identified by host cells and how it initiates the host antiviral defense. There are ten potential viral DNA sensors that have been reported. However, they have not been confirmed as consistently used sensors for distinctive DNA viruses in different animal and cell types [102Nie Y, Wang YY. Innate immune responses to DNA viruses. Protein Cell 2013; 4(1): 1-7.
[http://dx.doi.org/10.1007/s13238-012-2122-6] [PMID: 23307778] ].
Immune responses towards DNA are not limited to IFN-inducing pathways. Cytosolic DNA initiates caspase-1-dependent development of pro-inflammatory cytokines interleukin (IL)-18 and IL-1β. This pathway is facilitated by a Pyrin- and HIN200-domain-containing (PYHIN) protein called AIM2. Current data from knockout studies have shown the significance of AIM2 in host defense to DNA viruses. IFI16 is one of the most recent PYHIN proteins that have been acknowledged. IFI16 plays an important role in viral DNA attachment and the immune response of certain DNA viruses. Like AIM2, IFI16 joins viral DNA through HIN domains. IFI16 activation prompts inflammatory cytokine assembly and IFN-β in response to cytosolically administered HSV-1 or viral DNA infection [103Sharma S, Fitzgerald KA. Innate immune sensing of DNA. PLoS Pathog 2011; 7(4): e1001310.
[http://dx.doi.org/10.1371/journal.ppat.1001310] [PMID: 21533068] ].
Major pathogenic DNA viruses, such as human papillomavirus, polyoma virus, HSV, and EBV, have used many tactics to evade and weaken the immune response [104Whitley RJ. Herpesviruses. In: Baron S, Ed. Medical Microbiology 1996.]. Recent studies have shown that these viruses modify recipient host cell responses. This can lead to the formation of productive infections. In addition, many viruses obtain host genes. This is needed to transform them for a specific purpose to avoid targeting by the immune system. Controls of viral infections are often facilitated by the stimulation of innate and adaptive immune mechanisms that include several types of cells. B cells, CD4+ T cells, and CD8+ T cells contribute to anti-viral responses in an antigen-specific fashion. This contribution is done by antibody secretion, cytolytic effect, or IFN-γ production, to guarantee extended protection. Alternatively, the stromal cells, NK-cells, and antigen-presenting cells play a major role in virus control during the early phase of infection. Though type I IFNs are important in forming early anti-viral defenses, other cytokines, such as IL-1β, IL-18, and IL-12 contribute to early defense in anti-viral immunity [102Nie Y, Wang YY. Innate immune responses to DNA viruses. Protein Cell 2013; 4(1): 1-7.
[http://dx.doi.org/10.1007/s13238-012-2122-6] [PMID: 23307778] ].
10. ROLE OF EXOSOME BIOGENESIS IN IMMUNE RESPONSES TO VIRAL INFECTION
Host cell machinery utilizes vesicle secretion as a defense mechanism against viral infection [12Meckes DG Jr, Raab-Traub N. Microvesicles and viral infection. J Virol 2011; 85(24): 12844-54.
[http://dx.doi.org/10.1128/JVI.05853-11] [PMID: 21976651] ]. Secreted vesicles may stimulate immune cells and present viral antigens during a cellular response. For instance, the apolipoprotein B mRNA editing enzyme catalytic subunit 3G (APOBEC3G),-a cytidine deaminase that participates in the antiviral cellular network against retroviruses, can be packed and delivered to neighboring cells through exosomes to prevent HIV-1 replication [12Meckes DG Jr, Raab-Traub N. Microvesicles and viral infection. J Virol 2011; 85(24): 12844-54.
[http://dx.doi.org/10.1128/JVI.05853-11] [PMID: 21976651] ]. Results from Izquierdo Useros et al. showed that HIV-1 uses DCs as a transit location in the non-replicative stage [80Izquierdo-Useros N, Lorizate M, Puertas MC, et al. Siglec-1 is a novel dendritic cell receptor that mediates HIV-1 trans-infection through recognition of viral membrane gangliosides. PLoS Biol 2012; 10(12): e1001448.
[http://dx.doi.org/10.1371/journal.pbio.1001448] [PMID: 23271952] ]. The virus packages all viral antigens and particles in exosome-like vesicles after fusing with DCs [80Izquierdo-Useros N, Lorizate M, Puertas MC, et al. Siglec-1 is a novel dendritic cell receptor that mediates HIV-1 trans-infection through recognition of viral membrane gangliosides. PLoS Biol 2012; 10(12): e1001448.
[http://dx.doi.org/10.1371/journal.pbio.1001448] [PMID: 23271952] ]. Exosomes bearing viral antigens deliver their cargo to CD4+ T cells and provoke infection [40van Dongen HM, Masoumi N, Witwer KW, Pegtel DM. Extracellular vesicles exploit viral entry routes for cargo delivery. Microbiol Mol Biol Rev 2016; 80(2): 369-86.
[http://dx.doi.org/10.1128/MMBR.00063-15] [PMID: 26935137] ] (Table 1).
Exosomes can be used as therapeutic agents to modulate immune responses [105Robbins PD, Morelli AE. Regulation of immune responses by extracellular vesicles. Nat Rev Immunol 2014; 14(3): 195-208.
[http://dx.doi.org/10.1038/nri3622] [PMID: 24566916] ]. Exosomes mediate intercellular communication via innate and adaptive immune responses [56Alenquer M, Amorim MJ. Exosome Biogenesis, Regulation, and Function in Viral Infection. Viruses 2015; 7(9): 5066-83.
[http://dx.doi.org/10.3390/v7092862] [PMID: 26393640] ]. A variety of cells (B and T lymphocytes and dendritic cells) of the immune system have been shown to release exosome vesicles displaying immune modulatory properties [56Alenquer M, Amorim MJ. Exosome Biogenesis, Regulation, and Function in Viral Infection. Viruses 2015; 7(9): 5066-83.
[http://dx.doi.org/10.3390/v7092862] [PMID: 26393640] ].
The immunological properties of exosomes were first reported in 1996 by Raposo and her collaborators [90Raposo G, Nijman HW, Stoorvogel W, et al. B lymphocytes secrete antigen-presenting vesicles. J Exp Med 1996; 183(3): 1161-72.
[http://dx.doi.org/10.1084/jem.183.3.1161] [PMID: 8642258] ]. The group reported that B lymphocytes infected with EBV released exosomes that contained MHC II molecules that were essential to adaptive immune response [90Raposo G, Nijman HW, Stoorvogel W, et al. B lymphocytes secrete antigen-presenting vesicles. J Exp Med 1996; 183(3): 1161-72.
[http://dx.doi.org/10.1084/jem.183.3.1161] [PMID: 8642258] ]. In addition, they reported that these exosomes could activate specific CD4+ T cell clones in vitro, suggesting that they could play a role in adaptive immune responses [90Raposo G, Nijman HW, Stoorvogel W, et al. B lymphocytes secrete antigen-presenting vesicles. J Exp Med 1996; 183(3): 1161-72.
[http://dx.doi.org/10.1084/jem.183.3.1161] [PMID: 8642258] ]. In another study, Pegtel et al. showed that infected EBV derived exosomes deliver miRNA cargo [76Pegtel DM, Cosmopoulos K, Thorley-Lawson DA, et al. Functional delivery of viral miRNAs via exosomes. Proc Natl Acad Sci USA 2010; 107(14): 6328-33.
[http://dx.doi.org/10.1073/pnas.0914843107] [PMID: 20304794] ]. They showed that these exosomes could move through the bloodstream and affect healthy recipient cells [76Pegtel DM, Cosmopoulos K, Thorley-Lawson DA, et al. Functional delivery of viral miRNAs via exosomes. Proc Natl Acad Sci USA 2010; 107(14): 6328-33.
[http://dx.doi.org/10.1073/pnas.0914843107] [PMID: 20304794] ].
Exosomes can influence major tumor-related pathways [106Azmi AS, Bao B, Sarkar FH. Exosomes in cancer development, metastasis, and drug resistance: A comprehensive review. Cancer Metastasis Rev 2013; 32(3-4): 623-42.
[http://dx.doi.org/10.1007/s10555-013-9441-9] [PMID: 23709120] ]. Exosomes can alter tumor microenvironment by regulating metastasis, angiogenesis, and immunity [107Young Hwa Soung SF, Ford S, Zhang V, Chung J. Exosomes in cancer diagnostics. Cancers 2017; 9(1): 8.
[http://dx.doi.org/10.3390/cancers9010008] ]. The first study highlighting the role of exosomes in cancer was reported in 1996 by Zitvogel and his colleagues [108Zitvogel L, Regnault A, Lozier A, et al. Eradication of established murine tumors using a novel cell-free vaccine: Dendritic cell-derived exosomes. Nat Med 1998; 4(5): 594-600.
[http://dx.doi.org/10.1038/nm0598-594] [PMID: 9585234] ]. They reported that exosomes released by DCs could promote the induction of CD8+ T-lymphocyte dependent anti-tumor immune responses in mice in vivo (77). Because of these findings, they concluded that exosome-based cell-free vaccines could be used as an alternative method for DC adoptive therapy against tumors [106Azmi AS, Bao B, Sarkar FH. Exosomes in cancer development, metastasis, and drug resistance: A comprehensive review. Cancer Metastasis Rev 2013; 32(3-4): 623-42.
[http://dx.doi.org/10.1007/s10555-013-9441-9] [PMID: 23709120] ].
CONCLUSION
Exosomes are tiny sized vesicles derived from cells that facilitate intercellular communication processes. Numerous studies have demonstrated that virally infected cells alter the host exosomal composition (Table 1). Many viruses can hijack host exosome pathways. Modified exosomes contribute to viral transmission and immune invasion. This review highlights the role of exosomes as a vehicle/non-classical entry pathway for RNA and DNA viruses.
There have been numerous amounts of studies highlighting the relationship of RNA viruses, immune response and exosome regulation. However, the impact of DNA virus infections and their effect on exosome biogenesis have not yet been completely explained. Exosomes can help the host restrict and/or enhance immune responses against pathogens by activating anti-viral mechanisms. Currently, in our laboratory, we are evaluating exosome biogenesis and composition and their role(s) in DNA viral infection and/or host protection. Clarifying these mechanisms may lead to better diagnostics and alternative therapeutics against a variety of common and emerging viral infections.
CONSENT FOR PUBLICATION
Not applicable.
CONFLICT OF INTEREST
The authors declare no conflict of interest, financial or otherwise.
ACKNOWLEDGMENTS
This work was supported by the American Association of Immunologists Careers in Immunology Fellowship Program (to QLM), Gorgas Memorial Foundation Research Grant Award (QLM). National Institutes of Health grant #5R01AI089337-04, 1R15 DA045564-01 (to QLM), and University of Alabama at Birmingham (UAB) Center for AIDS Research (CFAR), an NIH-funded program (P30 AI027767) made possible by the following institutes: NIAID, NCI, NICHD, NHLBI, NIDA, NIMH, NIA, NIDDK, NIGMS, FIC, and OAR. We are thankful to the scientific support from UABs: Preclinical Imaging Shared Facility. UAB Comprehensive Cancer Center’s Preclinical Imaging Shared Facility (Center Core Support Grant P30 CA013148).
REFERENCES
| [1] | Chahar HS, Bao X, Casola A. Exosomes and their role in the life cycle and pathogenesis of RNA viruses. Viruses 2015; 7(6): 3204-25. [http://dx.doi.org/10.3390/v7062770] [PMID: 26102580] |
| [2] | Théry C. Exosomes: Secreted vesicles and intercellular communications. F1000 Biol Rep 2011; 3: 15. [http://dx.doi.org/10.3410/B3-15] [PMID: 21876726] |
| [3] | Raposo G, Stoorvogel W. Extracellular vesicles: Exosomes, microvesicles, and friends. J Cell Biol 2013; 200(4): 373-83. [http://dx.doi.org/10.1083/jcb.201211138] [PMID: 23420871] |
| [4] | van Niel G, Raposo G, Candalh C, et al. Intestinal epithelial cells secrete exosome-like vesicles. Gastroenterology 2001; 121(2): 337-49. [http://dx.doi.org/10.1053/gast.2001.26263] [PMID: 11487543] |
| [5] | Thery C, Amigorena S, Raposo G, et al. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr Protoc Cell Biol 2006. Chapter 3; Unit 3 22. [http://dx.doi.org/10.1002/0471143030.cb0322s30] [PMID: 18228490] |
| [6] | De Toro J, Herschlik L, Waldner C, Mongini C. Emerging roles of exosomes in normal and pathological conditions: new insights for diagnosis and therapeutic applications. Front Immunolo 2015; 6(203): 1-12. [http://dx.doi.org/10.3389/fimmu.2015.00203] [PMID: 25999947] |
| [7] | Dear JW, Street JM, Bailey MA. Urinary exosomes: A reservoir for biomarker discovery and potential mediators of intrarenal signalling. Proteomics 2013; 13(10-11): 1572-80. [http://dx.doi.org/10.1002/pmic.201200285] [PMID: 23129434] |
| [8] | Madison MN, Roller RJ, Okeoma CM. Human semen contains exosomes with potent anti-HIV-1 activity. Retrovirology 2014; 11: 102. [http://dx.doi.org/10.1186/s12977-014-0102-z] [PMID: 25407601] |
| [9] | Lässer C, Alikhani VS, Ekström K, et al. Human saliva, plasma and breast milk exosomes contain RNA: Uptake by macrophages. J Transl Med 2011; 9: 9. [http://dx.doi.org/10.1186/1479-5876-9-9] [PMID: 21235781] |
| [10] | Street JM, Barran PE, Mackay CL, et al. Identification and proteomic profiling of exosomes in human cerebrospinal fluid. J Transl Med 2012; 10: 5. [http://dx.doi.org/10.1186/1479-5876-10-5] [PMID: 22221959] |
| [11] | Näslund TI, Paquin-Proulx D, Paredes PT, Vallhov H, Sandberg JK, Gabrielsson S. Exosomes from breast milk inhibit HIV-1 infection of dendritic cells and subsequent viral transfer to CD4+ T cells. AIDS 2014; 28(2): 171-80. [http://dx.doi.org/10.1097/QAD.0000000000000159] [PMID: 24413309] |
| [12] | Meckes DG Jr, Raab-Traub N. Microvesicles and viral infection. J Virol 2011; 85(24): 12844-54. [http://dx.doi.org/10.1128/JVI.05853-11] [PMID: 21976651] |
| [13] | Colombo M, Moita C, van Niel G, et al. Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. J Cell Sci 2013; 126(Pt 24): 5553-65. [http://dx.doi.org/10.1242/jcs.128868] [PMID: 24105262] |
| [14] | Théry C, Zitvogel L, Amigorena S. Exosomes: Composition, biogenesis and function. Nat Rev Immunol 2002; 2(8): 569-79. [http://dx.doi.org/10.1038/nri855] [PMID: 12154376] |
| [15] | Pan BT, Johnstone RM. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: Selective externalization of the receptor. Cell 1983; 33(3): 967-78. [http://dx.doi.org/10.1016/0092-8674(83)90040-5] [PMID: 6307529] |
| [16] | Akers JC, Gonda D, Kim R, Carter BS, Chen CC. Biogenesis of Extracellular Vesicles (EV): Exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J Neurooncol 2013; 113(1): 1-11. [http://dx.doi.org/10.1007/s11060-013-1084-8] [PMID: 23456661] |
| [17] | Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J Biol Chem 1987; 262(19): 9412-20. [PMID: 3597417] |
| [18] | Kalra H, Drummen GP, Mathivanan S. Focus on extracellular vesicles: Introducing the next small big thing. Int J Mol Sci 2016; 17(2): 170. [http://dx.doi.org/10.3390/ijms17020170] [PMID: 26861301] |
| [19] | Beach A, Zhang HG, Ratajczak MZ, Kakar SS. Exosomes: An overview of biogenesis, composition and role in ovarian cancer. J Ovarian Res 2014; 7: 14. [http://dx.doi.org/10.1186/1757-2215-7-14] [PMID: 24460816] |
| [20] | Trajkovic K, Hsu C, Chiantia S, et al. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science 2008; 319(5867): 1244-7. [http://dx.doi.org/10.1126/science.1153124] [PMID: 18309083] |
| [21] | Bassereau P. Division of labour in ESCRT complexes. Nat Cell Biol 2010; 12(5): 422-3. [http://dx.doi.org/10.1038/ncb0510-422] [PMID: 20442701] |
| [22] | Teo H, Gill DJ, Sun J, et al. ESCRT-I core and ESCRT-II GLUE domain structures reveal role for GLUE in linking to ESCRT-I and membranes. Cell 2006; 125(1): 99-111. [http://dx.doi.org/10.1016/j.cell.2006.01.047] [PMID: 16615893] |
| [23] | Stahelin RV, Long F, Diraviyam K. Phosphatidylinositol 3-phosphate induces the membrane penetration of the FYVE domains of Vps27p and Hr. J Biol Chem 2002; 277(29): 26379-88. [http://dx.doi.org/10.1074/jbc.M201106200] [PMID: 12006563] |
| [24] | Stuffers S, Sem Wegner C, Stenmark H, Brech A. Multivesicular endosome biogenesis in the absence of ESCRTs. Traffic 2009; 10(7): 925-37. [http://dx.doi.org/10.1111/j.1600-0854.2009.00920.x] [PMID: 19490536] |
| [25] | Yuana Y, Sturk A, Nieuwland R. Extracellular vesicles in physiological and pathological conditions. Blood Rev 2013; 27(1): 31-9. [http://dx.doi.org/10.1016/j.blre.2012.12.002] [PMID: 23261067] |
| [26] | Beach A, Zhang HG, Ratajczak MZ, Kakar SS. Exosomes: An overview of biogenesis, composition and role in ovarian cancer. J Ovarian Res 2014; 7: 14-4. [http://dx.doi.org/10.1186/1757-2215-7-14] [PMID: 24460816] |
| [27] | Zhang J, Li S, Li L, et al. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genomics Proteomics Bioinformatics 2015; 13(1): 17-24. [http://dx.doi.org/10.1016/j.gpb.2015.02.001] [PMID: 25724326] |
| [28] | Ostrowski M, Carmo NB, Krumeich S, et al. Rab27a and Rab27b control different steps of the exosome secretion pathway. Nat Cell Biol 2010; 12(1): 19-30. [http://dx.doi.org/10.1038/ncb2000] [PMID: 19966785] |
| [29] | Baietti MF, Zhang Z, Mortier E, et al. Syndecan-syntenin-ALIX regulates the biogenesis of exosomes. Nat Cell Biol 2012; 14(7): 677-85. [http://dx.doi.org/10.1038/ncb2502] [PMID: 22660413] |
| [30] | Yu X, Harris SL, Levine AJ. The regulation of exosome secretion: A novel function of the p53 protein. Cancer Res 2006; 66(9): 4795-801. [http://dx.doi.org/10.1158/0008-5472.CAN-05-4579] [PMID: 16651434] |
| [31] | Greening DW, Xu R, Ji H, Tauro BJ, Simpson RJ. A protocol for exosome isolation and characterization: Evaluation of ultracentrifugation, density-gradient separation, and immunoaffinity capture methods. Methods Mol Biol 2015; 1295: 179-209. [http://dx.doi.org/10.1007/978-1-4939-2550-6_15] [PMID: 25820723] |
| [32] | Alvarez ML, Khosroheidari M, Kanchi Ravi R, DiStefano JK. Comparison of protein, microRNA, and mRNA yields using different methods of urinary exosome isolation for the discovery of kidney disease biomarkers. Kidney Int 2012; 82(9): 1024-32. [http://dx.doi.org/10.1038/ki.2012.256] [PMID: 22785172] |
| [33] | Tauro BJ, Greening DW, Mathias RA, et al. Comparison of ultracentrifugation, density gradient separation, and immunoaffinity capture methods for isolating human colon cancer cell line LIM1863-derived exosomes. Methods 2012; 56(2): 293-304. [http://dx.doi.org/10.1016/j.ymeth.2012.01.002] [PMID: 22285593] |
| [34] | Li P, Kaslan M, Lee SH, Yao J, Gao Z. Progress in exosome isolation techniques. Theranostics 2017; 7(3): 789-804. [http://dx.doi.org/10.7150/thno.18133] [PMID: 28255367] |
| [35] | Chen C, Skog J, Hsu CH, et al. Microfluidic isolation and transcriptome analysis of serum microvesicles. Lab Chip 2010; 10(4): 505-11. [http://dx.doi.org/10.1039/B916199F] [PMID: 20126692] |
| [36] | Zhang Y, Bergelson JM. Adenovirus receptors. J Virol 2005; 79(19): 12125-31. [http://dx.doi.org/10.1128/JVI.79.19.12125-12131.2005] [PMID: 16160140] |
| [37] | Konadu KA, Huang MB, Roth W, et al. Isolation of exosomes from the plasma of HIV-1 positive individuals. J Vis Exp 2016; 107: 53495. [http://dx.doi.org/10.3791/53495] [PMID: 26780239] |
| [38] | Koliha N, Wiencek Y, Heider U, et al. A novel multiplex bead-based platform highlights the diversity of extracellular vesicles. J Extracell Vesicles 2016; 5: 29975. [http://dx.doi.org/10.3402/jev.v5.29975] [PMID: 26901056] |
| [39] | Madison MN, Jones PH, Okeoma CM. Exosomes in human semen restrict HIV-1 transmission by vaginal cells and block intravaginal replication of LP-BM5 murine AIDS virus complex. Virology 2015; 482: 189-201. [http://dx.doi.org/10.1016/j.virol.2015.03.040] [PMID: 25880110] |
| [40] | van Dongen HM, Masoumi N, Witwer KW, Pegtel DM. Extracellular vesicles exploit viral entry routes for cargo delivery. Microbiol Mol Biol Rev 2016; 80(2): 369-86. [http://dx.doi.org/10.1128/MMBR.00063-15] [PMID: 26935137] |
| [41] | Longatti A. The dual role of exosomes in hepatitis A and C virus transmission and viral immune activation. Viruses 2015; 7(12): 6707-15. [http://dx.doi.org/10.3390/v7122967] [PMID: 26694453] |
| [42] | Anderson MR, Kashanchi F, Jacobson S. Exosomes in viral disease. Neurotherapeutics 2016; 13(3): 535-46. [http://dx.doi.org/10.1007/s13311-016-0450-6] [PMID: 27324390] |
| [43] | Ramakrishnaiah V, Thumann C, Fofana I, et al. Exosome-mediated transmission of hepatitis C virus between human hepatoma Huh7.5 cells. Proc Natl Acad Sci USA 2013; 110(32): 13109-13. [http://dx.doi.org/10.1073/pnas.1221899110] [PMID: 23878230] |
| [44] | Nour AM, Modis Y. Endosomal vesicles as vehicles for viral genomes. Trends Cell Biol 2014; 24(8): 449-54. [http://dx.doi.org/10.1016/j.tcb.2014.03.006] [PMID: 24746011] |
| [45] | Smit JM, Moesker B, Rodenhuis-Zybert I, Wilschut J. Flavivirus cell entry and membrane fusion. Viruses 2011; 3(2): 160-71. [http://dx.doi.org/10.3390/v3020160] [PMID: 22049308] |
| [46] | Hamel R, Dejarnac O, Wichit S, et al. Biology of Zika virus infection in human skin cells. J Virol 2015; 89(17): 8880-96. [http://dx.doi.org/10.1128/JVI.00354-15] [PMID: 26085147] |
| [47] | Liu Z, Zhang X, Yu Q, He JJ. Exosome-associated hepatitis C virus in cell cultures and patient plasma. Biochem Biophys Res Commun 2014; 455(3-4): 218-22. [http://dx.doi.org/10.1016/j.bbrc.2014.10.146] [PMID: 25449270] |
| [48] | Dreux M, Garaigorta U, Boyd B, et al. Short-range exosomal transfer of viral RNA from infected cells to plasmacytoid dendritic cells triggers innate immunity. Cell Host Microbe 2012; 12(4): 558-70. [http://dx.doi.org/10.1016/j.chom.2012.08.010] [PMID: 23084922] |
| [49] | Lodish H, Berk A, Zipursky SL, et al. Viruses: structure, function, and uses. Molecular cell biology. (4th ed). New York, NY: W. H. Freeman 2000. |
| [50] | Byland R, Marsh M. Trafficking of viral membrane proteins. Curr Top Microbiol Immunol 2005; 285: 219-54. [http://dx.doi.org/10.1007/3-540-26764-6_8] [PMID: 15609506] |
| [51] | Munich S, Sobo-Vujanovic A, Buchser WJ, Beer-Stolz D, Vujanovic NL. Dendritic cell exosomes directly kill tumor cells and activate natural killer cells via TNF superfamily ligands. OncoImmunology 2012; 1(7): 1074-83. [http://dx.doi.org/10.4161/onci.20897] [PMID: 23170255] |
| [52] | Mulcahy LA, Pink RC, Carter DR. Routes and mechanisms of extracellular vesicle uptake. J Extracell Vesicles 2014; 3: 3. [http://dx.doi.org/10.3402/jev.v3.24641] [PMID: 25143819] |
| [53] | Tian T, Zhu YL, Hu FH, Wang YY, Huang NP, Xiao ZD. Dynamics of exosome internalization and trafficking. J Cell Physiol 2013; 228(7): 1487-95. [http://dx.doi.org/10.1002/jcp.24304] [PMID: 23254476] |
| [54] | Koumangoye RB, Sakwe AM, Goodwin JS, Patel T, Ochieng J. Detachment of breast tumor cells induces rapid secretion of exosomes which subsequently mediate cellular adhesion and spreading. PLoS One 2011; 6(9): e24234. [http://dx.doi.org/10.1371/journal.pone.0024234] [PMID: 21915303] |
| [55] | Escrevente C, Keller S, Altevogt P, Costa J. Interaction and uptake of exosomes by ovarian cancer cells. BMC Cancer 2011; 11: 108. [http://dx.doi.org/10.1186/1471-2407-11-108] [PMID: 21439085] |
| [56] | Alenquer M, Amorim MJ. Exosome Biogenesis, Regulation, and Function in Viral Infection. Viruses 2015; 7(9): 5066-83. [http://dx.doi.org/10.3390/v7092862] [PMID: 26393640] |
| [57] | Pelchen-Matthews A, Raposo G, Marsh M. Endosomes, exosomes and Trojan viruses. Trends Microbiol 2004; 12(7): 310-6. [http://dx.doi.org/10.1016/j.tim.2004.05.004] [PMID: 15223058] |
| [58] | Hu WS, Hughes SH. HIV-1 reverse transcription. Cold Spring Harb Perspect Med 2012; 2(10): a006882. [http://dx.doi.org/10.1101/cshperspect.a006882] [PMID: 23028129] |
| [59] | Coffin JM, Hughes SH, Varmus HE. The interactions of retroviruses and their hosts.Retroviruses 1997. |
| [60] | Anson DS. The use of retroviral vectors for gene therapy-what are the risks? A review of retroviral pathogenesis and its relevance to retroviral vector-mediated gene delivery. Genet Vaccines Ther 2004; 2(1): 9. [http://dx.doi.org/10.1186/1479-0556-2-9] [PMID: 15310406] |
| [61] | Izquierdo-Useros N, Puertas MC, Borràs FE, Blanco J, Martinez-Picado J. Exosomes and retroviruses: The chicken or the egg? Cell Microbiol 2011; 13(1): 10-7. [http://dx.doi.org/10.1111/j.1462-5822.2010.01542.x] [PMID: 21054740] |
| [62] | Sharp PM, Hahn BH. Origins of HIV and the AIDS pandemic. Cold Spring Harb Perspect Med 2011; 1(1): a006841. [http://dx.doi.org/10.1101/cshperspect.a006841] [PMID: 22229120] |
| [63] | Blut A. Human Immunodeficiency Virus (HIV). Transfus Med Hemother 2016; 43(3): 203-22. [http://dx.doi.org/10.1159/000445852] [PMID: 27403093] |
| [64] | Cohen MS, Hellmann N, Levy JA, DeCock K, Lange J. The spread, treatment, and prevention of HIV-1: Evolution of a global pandemic. J Clin Invest 2008; 118(4): 1244-54. [http://dx.doi.org/10.1172/JCI34706] [PMID: 18382737] |
| [65] | de Carvalho JV, de Castro RO, da Silva EZ, et al. Nef neutralizes the ability of exosomes from CD4+ T cells to act as decoys during HIV-1 infection. PLoS One 2014; 9(11): e113691. [http://dx.doi.org/10.1371/journal.pone.0113691] [PMID: 25423108] |
| [66] | Ray N, Doms RW. HIV-1 coreceptors and their inhibitors. Curr Top Microbiol Immunol 2006; 303: 97-120. [http://dx.doi.org/10.1007/978-3-540-33397-5_5] [PMID: 16570858] |
| [67] | Melikyan GB. HIV entry: A game of hide-and-fuse? Curr Opin Virol 2014; 4: 1-7. [http://dx.doi.org/10.1016/j.coviro.2013.09.004] [PMID: 24525288] |
| [68] | Benson RE, Sanfridson A, Ottinger JS, Doyle C, Cullen BR. Downregulation of cell-surface CD4 expression by simian immunodeficiency virus Nef prevents viral super infection. J Exp Med 1993; 177(6): 1561-6. [http://dx.doi.org/10.1084/jem.177.6.1561] [PMID: 8098729] |
| [69] | Kowal J, Tkach M, Théry C. Biogenesis and secretion of exosomes. Curr Opin Cell Biol 2014; 29: 116-25. [http://dx.doi.org/10.1016/j.ceb.2014.05.004] [PMID: 24959705] |
| [70] | Conde-Vancells J, Rodriguez-Suarez E, Embade N, et al. Characterization and comprehensive proteome profiling of exosomes secreted by hepatocytes. J Proteome Res 2008; 7(12): 5157-66. [http://dx.doi.org/10.1021/pr8004887] [PMID: 19367702] |
| [71] | Théry C, Boussac M, Véron P, et al. Proteomic analysis of dendritic cell-derived exosomes: A secreted subcellular compartment distinct from apoptotic vesicles. J Immunol 2001; 166(12): 7309-18. [http://dx.doi.org/10.4049/jimmunol.166.12.7309] [PMID: 11390481] |
| [72] | Krishnamoorthy L, Bess JW Jr, Preston AB, Nagashima K, Mahal LK. HIV-1 and microvesicles from T cells share a common glycome, arguing for a common origin. Nat Chem Biol 2009; 5(4): 244-50. [http://dx.doi.org/10.1038/nchembio.151] [PMID: 19234452] |
| [73] | Wubbolts R, Leckie RS, Veenhuizen PT, et al. Proteomic and biochemical analyses of human B cell-derived exosomes. Potential implications for their function and multivesicular body formation. J Biol Chem 2003; 278(13): 10963-72. [http://dx.doi.org/10.1074/jbc.M207550200] [PMID: 12519789] |
| [74] | Ott DE. Cellular proteins detected in HIV-1. Rev Med Virol 2008; 18(3): 159-75. [http://dx.doi.org/10.1002/rmv.570] [PMID: 18265424] |
| [75] | Valadi H, Ekström K, Bossios A, Sjöstrand M, Lee JJ, Lötvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol 2007; 9(6): 654-9. [http://dx.doi.org/10.1038/ncb1596] [PMID: 17486113] |
| [76] | Pegtel DM, Cosmopoulos K, Thorley-Lawson DA, et al. Functional delivery of viral miRNAs via exosomes. Proc Natl Acad Sci USA 2010; 107(14): 6328-33. [http://dx.doi.org/10.1073/pnas.0914843107] [PMID: 20304794] |
| [77] | Lenassi M, Cagney G, Liao M, et al. HIV Nef is secreted in exosomes and triggers apoptosis in bystander CD4+ T cells. Traffic 2010; 11(1): 110-22. [http://dx.doi.org/10.1111/j.1600-0854.2009.01006.x] [PMID: 19912576] |
| [78] | Grigorov B, Attuil-Audenis V, Perugi F, et al. A role for CD81 on the late steps of HIV-1 replication in a chronically infected T cell line. Retrovirology 2009; 6: 28. [http://dx.doi.org/10.1186/1742-4690-6-28] [PMID: 19284574] |
| [79] | Raymond AD, Campbell-Sims TC, Khan M, et al. HIV Type 1 Nef is released from infected cells in CD45(+) microvesicles and is present in the plasma of HIV-infected individuals. AIDS Res Hum Retroviruses 2011; 27(2): 167-78. [http://dx.doi.org/10.1089/aid.2009.0170] [PMID: 20964480] |
| [80] | Izquierdo-Useros N, Lorizate M, Puertas MC, et al. Siglec-1 is a novel dendritic cell receptor that mediates HIV-1 trans-infection through recognition of viral membrane gangliosides. PLoS Biol 2012; 10(12): e1001448. [http://dx.doi.org/10.1371/journal.pbio.1001448] [PMID: 23271952] |
| [81] | Madison MN, Okeoma CM. Exosomes: Implications in HIV-1 Pathogenesis. Viruses 2015; 7(7): 4093-118. [http://dx.doi.org/10.3390/v7072810] [PMID: 26205405] |
| [82] | Roeth JF, Williams M, Kasper MR, Filzen TM, Collins KL. HIV-1 Nef disrupts MHC-I trafficking by recruiting AP-1 to the MHC-I cytoplasmic tail. J Cell Biol 2004; 167(5): 903-13. [http://dx.doi.org/10.1083/jcb.200407031] [PMID: 15569716] |
| [83] | Chaudhuri R, Mattera R, Lindwasser OW, Robinson MS, Bonifacino JS. A basic patch on alpha-adaptin is required for binding of human immunodeficiency virus type 1 Nef and cooperative assembly of a CD4-Nef-AP-2 complex. J Virol 2009; 83(6): 2518-30. [http://dx.doi.org/10.1128/JVI.02227-08] [PMID: 19129443] |
| [84] | Hung CH, Thomas L, Ruby CE, et al. HIV-1 Nef assembles a Src family kinase-ZAP-70/Syk-PI3K cascade to downregulate cell-surface MHC-I. Cell Host Microbe 2007; 1(2): 121-33. [http://dx.doi.org/10.1016/j.chom.2007.03.004] [PMID: 18005690] |
| [85] | Lubben NB, Sahlender DA, Motley AM, Lehner PJ, Benaroch P, Robinson MS. HIV-1 Nef-induced down-regulation of MHC class I requires AP-1 and clathrin but not PACS-1 and is impeded by AP-2. Mol Biol Cell 2007; 18(9): 3351-65. [http://dx.doi.org/10.1091/mbc.e07-03-0218] [PMID: 17581864] |
| [86] | Sims B, Farrow AL, Williams SD, et al. Role of TIM-4 in exosome-dependent entry of HIV-1 into human immune cells. Int J Nanome 2017; 12: 4823-33. [http://dx.doi.org/10.2147/IJN.S132762] [PMID: 28740388] |
| [87] | Jemielity S, Wang JJ, Chan YK, et al. TIM-family proteins promote infection of multiple enveloped viruses through virion-associated phosphatidylserine. PLoS Pathog 2013; 9(3): e1003232. [http://dx.doi.org/10.1371/journal.ppat.1003232] [PMID: 23555248] |
| [88] | Wilen CB, Tilton JC, Doms RW. HIV: Cell binding and entry. Cold Spring Harb Perspect Med 2012; 2(8): a006866. [http://dx.doi.org/10.1101/cshperspect.a006866] [PMID: 22908191] |
| [89] | Arenaccio C, Chiozzini C, Columba-Cabezas S, et al. Exosomes from human immunodeficiency virus type 1 (HIV-1)-infected cells license quiescent CD4+ T lymphocytes to replicate HIV-1 through a Nef- and ADAM17-dependent mechanism. J Virol 2014; 88(19): 11529-39. [http://dx.doi.org/10.1128/JVI.01712-14] [PMID: 25056899] |
| [90] | Raposo G, Nijman HW, Stoorvogel W, et al. B lymphocytes secrete antigen-presenting vesicles. J Exp Med 1996; 183(3): 1161-72. [http://dx.doi.org/10.1084/jem.183.3.1161] [PMID: 8642258] |
| [91] | Théry C, Regnault A, Garin J, et al. Molecular characterization of dendritic cell-derived exosomes. Selective accumulation of the heat shock protein hsc73. J Cell Biol 1999; 147(3): 599-610. [http://dx.doi.org/10.1083/jcb.147.3.599] [PMID: 10545503] |
| [92] | Raposo G, Tenza D, Mecheri S, Peronet R, Bonnerot C, Desaymard C. Accumulation of major histocompatibility complex class II molecules in mast cell secretory granules and their release upon degranulation. Mol Biol Cell 1997; 8(12): 2631-45. [http://dx.doi.org/10.1091/mbc.8.12.2631] [PMID: 9398681] |
| [93] | Blanchard N, Lankar D, Faure F, et al. TCR activation of human T cells induces the production of exosomes bearing the TCR/CD3/zeta complex. J Immunol 2002; 168(7): 3235-41. [http://dx.doi.org/10.4049/jimmunol.168.7.3235] [PMID: 11907077] |
| [94] | Martínez-Lorenzo MJ, Anel A, Gamen S, et al. Activated human T cells release bioactive Fas ligand and APO2 ligand in microvesicles. J Immunol 1999; 163(3): 1274-81. [PMID: 10415024] |
| [95] | Soares H. HIV-1 Intersection with CD4 T Cell Vesicle Exocytosis: Intercellular Communication Goes Viral. Front Immunol 2014; 5: 454. [http://dx.doi.org/10.3389/fimmu.2014.00454] [PMID: 25295039] |
| [96] | Sims B, Farrow AL, Williams SD, Bansal A, Krendelchtchikov A, Matthews QL. Tetraspanin blockage reduces exosome-mediated HIV-1 entry. Arch Virol 2018; 163(6): 1683-3. [http://dx.doi.org/10.1007/s00705-018-3737-6] [PMID: 29429034] |
| [97] | Gu L, Sims B, Krendelchtchikov A, Tabengwa E, Matthews QL. Differential binding of the HIV-1 envelope to phosphatidylserine receptors. Biochim Biophys Acta 2017; 1859(10): 1962-6. [http://dx.doi.org/10.1016/j.bbamem.2017.06.007] [PMID: 28622976] |
| [98] | Choi KH. Viral polymerases. Adv Exp Med Biol 2012; 726: 267-304. [http://dx.doi.org/10.1007/978-1-4614-0980-9_12] [PMID: 22297518] |
| [99] | Knipe DM. The role of viral and cellular nuclear proteins in herpes simplex virus replication. Adv Virus Res 1989; 37: 85-123. [http://dx.doi.org/10.1016/S0065-3527(08)60833-7] [PMID: 2557760] |
| [100] | Sims B, Gu L, Krendelchtchikov A, Matthews QL. Neural stem cell-derived exosomes mediate viral entry. Int J Nanomed 2014; 9: 4893-7. [http://dx.doi.org/10.2147/IJN.S70999] [PMID: 25364247] |
| [101] | Chaurushiya MS, Lilley CE, Aslanian A, et al. Viral E3 ubiquitin ligase-mediated degradation of a cellular E3: Viral mimicry of a cellular phosphorylation mark targets the RNF8 FHA domain. Mol Cell 2012; 46(1): 79-90. [http://dx.doi.org/10.1016/j.molcel.2012.02.004] [PMID: 22405594] |
| [102] | Nie Y, Wang YY. Innate immune responses to DNA viruses. Protein Cell 2013; 4(1): 1-7. [http://dx.doi.org/10.1007/s13238-012-2122-6] [PMID: 23307778] |
| [103] | Sharma S, Fitzgerald KA. Innate immune sensing of DNA. PLoS Pathog 2011; 7(4): e1001310. [http://dx.doi.org/10.1371/journal.ppat.1001310] [PMID: 21533068] |
| [104] | Whitley RJ. Herpesviruses. In: Baron S, Ed. Medical Microbiology 1996. |
| [105] | Robbins PD, Morelli AE. Regulation of immune responses by extracellular vesicles. Nat Rev Immunol 2014; 14(3): 195-208. [http://dx.doi.org/10.1038/nri3622] [PMID: 24566916] |
| [106] | Azmi AS, Bao B, Sarkar FH. Exosomes in cancer development, metastasis, and drug resistance: A comprehensive review. Cancer Metastasis Rev 2013; 32(3-4): 623-42. [http://dx.doi.org/10.1007/s10555-013-9441-9] [PMID: 23709120] |
| [107] | Young Hwa Soung SF, Ford S, Zhang V, Chung J. Exosomes in cancer diagnostics. Cancers 2017; 9(1): 8. [http://dx.doi.org/10.3390/cancers9010008] |
| [108] | Zitvogel L, Regnault A, Lozier A, et al. Eradication of established murine tumors using a novel cell-free vaccine: Dendritic cell-derived exosomes. Nat Med 1998; 4(5): 594-600. [http://dx.doi.org/10.1038/nm0598-594] [PMID: 9585234] |
| [109] | Guenat D, Hermetet F, Prétet JL, Mougin C. Exosomes and Other Extracellular Vesicles in HPV Transmission and Carcinogenesis. Viruses 2017; 9(8): E211. [http://dx.doi.org/10.3390/v9080211] [PMID: 28783104] |
| [110] | Kouwaki T, Fukushima Y, Daito T, et al. Extracellular vesicles including exosomes regulate innate immune responses to hepatitis B virus infection. Front Immunol 2016; 7(335) [http://dx.doi.org/10.3389/fimmu.2016.00335] |
| [111] | Abels ER, Breakefield XO. Introduction to Extracellular Vesicles: Biogenesis, RNA cargo selection, content, release, and uptake. Cell Mol Neurobiol 2016; 36(3): 301-12. [http://dx.doi.org/10.1007/s10571-016-0366-z] [PMID: 27053351] |
Endorsements
Browse Contents
Table of Contents
- INTRODUCTION
- HIJACKING BY VIRUSES EXPLOITS THE EXOSOMAL PATHWAY
- EXOSOMES AND PROTEIN TRAFFICKING
- EXOSOMES AND VIRAL INFECTIONS
- RETROVIRUSES
- HUMAN IMMUNODEFICIENCY VIRUS
- EXOSOMES AUGMENT HIV-1 ENTRY
- DNA VIRUSES HIJACKING
- IMMUNE RESPONSE TO DNA VIRUSES
- ROLE OF EXOSOME BIOGENESIS IN IMMUNE RESPONSES TO VIRAL INFECTION
- CONCLUSION