- Home
- About Journals
-
Information for Authors/ReviewersEditorial Policies
Publication Fee
Publication Cycle - Process Flowchart
Online Manuscript Submission and Tracking System
Publishing Ethics and Rectitude
Authorship
Author Benefits
Reviewer Guidelines
Guest Editor Guidelines
Peer Review Workflow
Quick Track Option
Copyediting Services
Bentham Open Membership
Bentham Open Advisory Board
Archiving Policies
Fabricating and Stating False Information
Post Publication Discussions and Corrections
Editorial Management
Advertise With Us
Funding Agencies
Rate List
Kudos
General FAQs
Special Fee Waivers and Discounts
- Contact
- Help
- About Us
- Search




Current Chemical Genomics and Translational Medicine
(Discontinued)
ISSN: 2213-9885 ― Volume 12, 2018
In Vitro Viability and Cytotoxicity Testing and Same-Well Multi-Parametric Combinations for High Throughput Screening
Andrew L. Niles*, Richard A. Moravec, Terry L. Riss
Abstract
In vitro cytotoxicity testing has become an integral aspect of drug discovery because it is a convenient, costeffective, and predictive means of characterizing the toxic potential of new chemical entities. The early and routine implementation of this testing is testament to its prognostic importance for humans. Although a plethora of assay chemistries and methods exist for 96-well formats, few are practical and sufficiently sensitive enough for application in high throughput screening (HTS). Here we briefly describe a handful of the currently most robust and validated HTS assays for accurate and efficient assessment of cytotoxic risk. We also provide guidance for successful HTS implementation and discuss unique merits and detractions inherent in each method. Lastly, we discuss the advantages of combining specific HTS compatible assays into multi-parametric, same-well formats.
Article Information
Identifiers and Pagination:
Year: 2009Volume: 3
First Page: 33
Last Page: 41
Publisher Id: CCGTM-3-33
DOI: 10.2174/1875397300903010033
Article History:
Received Date: 2/3/2009Revision Received Date: 4/4/2009
Acceptance Date: 17/4/2009
Electronic publication date: 11/6/2009
Collection year: 2009
open-access license: This is an open access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.5/), which permits unrestrictive use, distribution, and reproduction in any medium, provided the original work is properly cited.
* Address correspondence to this author at the Research Department, Promega Corporation, 2800 Woods Hollow Road, Madison, WI, USA; E-mail: andrew.niles@promega.com
| Open Peer Review Details | |||
|---|---|---|---|
| Manuscript submitted on 2-3-2009 |
Original Manuscript | In Vitro Viability and Cytotoxicity Testing and Same-Well Multi-Parametric Combinations for High Throughput Screening | |
INTRODUCTION
A significant advance in the field of early medicine was Paracelsus’ recognition that all compounds have the capacity to be poisonous depending upon dosage [1Rozman K, Doull J. Paracelsus, haber and arndt Toxicology 2001; 160: 191-96.]. This observation makes toxicity testing a necessary and critical industry practice to identify and define safety thresholds for all new potential chemotherapeutics. The advent of in vitro cytotoxicity testing has greatly streamlined this process and is now considered to be a nearly compulsory activity starting at target validation and continuing through medicinal modification.
Unlike animal-based toxicology testing, there are clearer definitions and greater agreement for what constitutes cytotoxicity in vitro. Classically speaking, a compound or treatment is considered to be cytotoxic if it prevents cellular attachment, causes dramatic morphological changes, adversely affects replication rate, or leads to a reduction in overall viability [2Horvath S. Cytotoxicity of drugs and diverse chemical agents to cell cultures Toxicology 1980; 16: 59-66.]. It should be noted that the manifestation of these effects is greatly dependent on length of compound exposure and mechanism of cytotoxicity [3Riss T, Moravec R. Use of multiple assay endpoints to investigate the effects of incubation time, dose of toxin, and plating density in cell-based cytotoxicity assays Assay Drug Dev Technol 2004; 2: 51-62.].
A host of new assays have been described and utilized which measure biomarkers of cellular stress or specific signaling events more proximal to initial cytotoxic insult (i.e. glutathione, caspases) [4Slim R, Toborek M, Robertson L, et al. Cellular glutathione status modulates polychlorinated biphenyl-induced status and apoptosis in vascular endothelial cells Toxicol Appl Pharm 2000; 166(1): 36-42.]. These methods offer early indication of potential cytotoxicity, but are typically relegated to secondary screening because they are more difficult to employ as endpoint assays due to the transient nature of the biomarker and kinetic differences associated with cell death progression [5Svingen PA, Loegering D, Rodriguez J, et al. Components of the cell death machine and drug sensitivity of the National Cancer Institute Cell Line Panel Clin Cancer Rev 2004; 10: 6807-20., 6Slee EA, Harte MT, Cluck RM, et al. Ordering the Cytochrome c-intitated caspase cascade: heirarchial activation of caspases-2,-3,-6,-7,-8, and -10 in a caspase-9 dependent manner J Cell Biol 1999; 144(2): 281-92.]. Therefore assay chemistries predicated upon the detection of changes in membrane integrity remain the gold standard for in vitro cytotoxicity testing.
Many methods exist for the assessment of membrane integrity, including several classic dye inclusion, exclusion and lysosomal accumulation techniques [7Cook J, Mitchell J. Viability measurements in mammalian cell systems Anal Biochem 1989; 179: 1-7., 8Borenfreund E, Puerner J. Toxicity determined in vitro by morphological alterations and neutral red adsorption Toxicol Lett 1985; 24: 119-24.]. Although well validated, these methods are poorly suited for HTS implementation due to low sensitivity, multiple processing steps or miniaturization problems associated with higher plate densities [9Rose D, Lemmo T. Challenges in implementing high density formats for high throughput screening Lab Autom News 1997; 2: 12-9., 10Maffia A, Kariv I, Oldenburg K. Miniaturization of a mammalian cell-based assay: luciferase reporter gene readout in a 3 microliter 1536-well plate J Biomol Screen 1999; 4: 137-42.]. Recent advances in reagent formulations accommodate these miniaturized formats and are fully compatible with automated dispensing systems and integrated detection instruments. These reagents deliver the linearity, sensitivity, and robustness (Z’ Values) necessary for properly interrogating large chemical libraries for cytotoxic risk. The most acceptable assay format is known as “add-mix-measure”, whereby reagent chemistry is delivered directly to the test well.
VIABILITY ASSAYS
Viability assays are designed to measure activities attributable to cellular maintenance and survival. These activities are typically metabolic biomarkers such as ATP and mitochondrial reductase potential, but can include homeostatic “housekeeping” enzyme activities. The underlying assay premise is that these activities are directly proportional to viable cell number after a treatment period. During relatively short incubation periods (8 hr or less), a reduction or complete cessation of these biomarker activities is strongly indicative of overt cytotoxicity by catastrophic membrane damage (i.e. primary necrosis) [11Nicotera P, Leist M, Manzo L. Neuronal cell death: a demise with different shapes Trends Pharmacol Sci 1999; 20: 46-51.]. A reduction in biomarker activity compared to control in longer incubations indicates either a reduction in normal cellular division rate (cell-cycle arrest) or cell death by programmed elimination mechanisms such as apoptosis [12Rixe O, Fojo T. Is cell death a critical end point for anticancer therapies or is cytostasis sufficient? Clin Cancer Res 2007; 13(24): 7280-87., 13Shi W, Gould MN. Induction of cytostasis in mammalian carcinoma cells treated with the anticancer agent perillyl alcohol Carcinogenesis 2002; 23: 131-42.]. Because viability assays measure the relative number of cells remaining after treatment, they offer significant utility even during long compound-contact periods (72 hr).
RESAZURIN REDUCTION
The reductive capacity inherent within viable cells can be conveniently measured using the redox indicator, resazurin [14O’Brien J, Wilson I, Orton T, et al. Investigation of the Alamar Blue (resazurin) fluorescent dye for the assessment of mammalian cell cytotoxicity Eur J Biochem 2000; 267: 5421-26., 15Page B, Page M, Noel C. A new fluorometric assay for cytotoxicity measurements in vitro Int J Oncol 1993; 3: 473-76.]. Resazurin is soluble in physiologically buffered formulations and can be added directly to growing cultures in a one-step, homogeneous addition. Unlike the tetrazolium chemistries (2,3-bis(2-methoxy-4-nitro-5-sulphophenyl)-5-carboxanilide-2H-tetrazolium (XTT), (4-[3-4-iodophenyl]-2-(4-nitrophenyl)-2H-5-tetrazolio)-1,3-benzene disulfonate) WST-1, 5-(3-carboxymethoxyphenyl)-2-(4, 5-dimethyl-thiazoly)-3-(4-sulfophenyl) tetrazolium, inner salt (MTS) which measure reductive capacity by colorimetric means, the product of resazurin reduction (resorufin) can also be measured using fluorescence with a fluorometer equipped with 560nm excitation and 590nm emission filter set [16Roehm N, Rodgers G, Hatfield S, et al. An improved colorimetric assay for cell proliferation and viability utilizing the tetrazolium salt XTT J Immunol Methods 1991; 142: 257-65.-18Cory A, Owen T, Barltrop J, et al. Use of an aqueous soluble tetrazolium/formazan assay for cell growth assays in culture Cancer Commun 1991; 3: 207-12.].
Resorufin product formation as a result of reagent contact with viable cells is accumulative and proceeds as a function of time. Although somewhat dependent upon cell type, metabolic rate and number per well, 2-4 hr of incubation with the reagent is typically sufficient to generate signal windows for adequate statistical discrimination in HTS. (Table 1) Fluorescent resazurin reductase assay chemistries have been adapted to 1536 well formats, but are typically most useful at lesser plate densities (384 and 96) due to practical limitations associated with prolonged incubation periods [19Shum D, Radu C, Kim E, et al. A high density assay format for he detection of novel cytotoxic agents in large chemical libraries J Enz Inhib Med Chem 2008; 23(6): 931-45., 20Hamid R, Rotshteyn Y, Rabadi L, et al. Comparison of alamar blue and MTT assays for high through-put screening Toxicol In Vitro 2004; 18: 703-10.].

Resazurin-based chemistries continue to offer significant value for HTS because the reagents are robust and stable, well-validated, and easy to use. The single greatest attribute however, is that resazurin reagents generate amongst the most cost-effective data on a per well basis [21Miret S, De Groene E, Klaffke W. Comparison of in vitro assays of cellular toxicity in the human hepatic cell line HepG2 J Biol Screen 2006; 11: 184-93.]. The major disadvantages of employing the chemistry are that the reagent is subject to interferences from compounds with inherent reductive capacity (such as ascorbic acid, glutathione, coenzyme A, dithiothreitol etc.) or those with intrinsic fluorescence characteristics [22Ulukaya E, Ozdikicioglu F, Yilmaztepe-Oral A, et al. The MTT assay yields a relatively lower result of growth inhibition than the ATP assay depending on the chemotherapeutic drugs tested Toxicol In Vitro 2008; 22: 232-39.-24Collier A, Pritsos C. The mitochondrial uncoupler dicumerol disrupts the MTT assay Biochem Pharm 2003; 66: 281-87.]. Lastly, extended reagent incubations (2hr or longer) may produce toxic affects or augment affects produced from the experimental compound [25Squatrito R, Connor J, Buller R. Comparison of a novel redox dye cell growth assay to the ATP bioluminescence assay Gynecol Oncol 1995; 58: 101-05.].
AMINOPEPTIDASE(S)
Eukaryotic cells contain a diverse abundance of proteolytic activities which act in a concerted effort to maintain homeostasis. Recently, a constitutive and conserved aminopeptidase-like, proteolytic biomarker profile was identified in mammalian cells that can be harnessed for assessing viability in cell culture [26Niles A, Moravec R, Hesselbeth P, et al. A homogeneous assay to measure live and dead cells in the same sample by detecting different protease markers Anal Biochem 2007; 366: 197-206.]. The assay utilizes the cell-permeant protease substrate glycyl-phenylalanyl-amino-fluorocoumerin (GF-AFC) that is delivered to wells in a neutral buffer. The substrate, which lacks an amino-terminal blocking moiety (i.e. Boc, Ac, or Cbz), can be processed by aminopeptidase(s) within the cytoplasm. The AFC product released after proteolysis of the substrate is proportional to viable cell number. Viability can be measured using a fluorometer equipped with a 380-400nm excitation source and a 505nm emission filter.
The selective detection of viable cells by this method is possible because the proteolytic activity towards the GF-AFC substrate is dependent upon the continued maintenance of membrane integrity. This live-cell, proteolytic activity decays within seconds after a cytotoxic event, so non-viable cells do not contribute appreciably to fluorescence generation. A 30 minute incubation of reagent and cells at 37ºC is typically sufficient for generating a workable signal window, but the incubation can be extended without detriment if work-flow scheduling does not permit shorter periods.
A major advantage of this assay is that it can be conveniently multiplexed with other compatible assay chemistries to deliver more per-well information (Fig. 1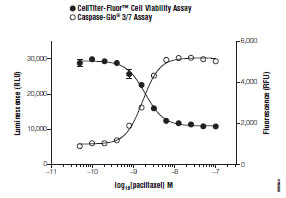 ) [27Niles A, Moravec R, Riss T. Update on in vitro cytotoxicity assays for drug development Expert Opin Drug Discov 2008; 3(6): 655-9.]. This feature is particularly attractive for normalizing spectrally-distinct fluorescent or luminescent data sets such as genetic report assays [28Niles A, Worzela T, Scurria M, et al. Multiplexed viability, cytotoxicity and apoptosis assays for cell-based screening Cell Notes 2006; 16: 12-5.]. Multiplex applications for viability and cytotoxicity will be handled in greater detail in a later section of this review. Lastly, although red-shifted from much of the problematic coumarin spectrum, the assay is susceptible to fluorescence interferences from test compounds [29Turek-Etienne T, Lei M, Terracciano J, et al. Use of red-shifted dyes in a fluorescence polarization AKT kinase assay for detection of biological activity in natural product extracts J Biomol Screen 2004; 9: 52-61.]. Additional interferences should be expected from protease inhibitors or from color quenching chemical entities.
) [27Niles A, Moravec R, Riss T. Update on in vitro cytotoxicity assays for drug development Expert Opin Drug Discov 2008; 3(6): 655-9.]. This feature is particularly attractive for normalizing spectrally-distinct fluorescent or luminescent data sets such as genetic report assays [28Niles A, Worzela T, Scurria M, et al. Multiplexed viability, cytotoxicity and apoptosis assays for cell-based screening Cell Notes 2006; 16: 12-5.]. Multiplex applications for viability and cytotoxicity will be handled in greater detail in a later section of this review. Lastly, although red-shifted from much of the problematic coumarin spectrum, the assay is susceptible to fluorescence interferences from test compounds [29Turek-Etienne T, Lei M, Terracciano J, et al. Use of red-shifted dyes in a fluorescence polarization AKT kinase assay for detection of biological activity in natural product extracts J Biomol Screen 2004; 9: 52-61.]. Additional interferences should be expected from protease inhibitors or from color quenching chemical entities.

ATP
ATP quantification is a widely accepted method for assessing viability because healthy cells contain closely regulated levels of the biomarker [30Crouch S, Kozlowski R, Slater K, et al. The use of ATP bioluminescence as a measure of cell proliferation and cytotoxicity J Immunol Methods 1993; 160: 81-., 31Lundin A. Estimation of biomass in growing cell lines by ATP assay Methods Enzymol 1986; 133: 27-42.]. Non-viable cells not only lose the ability to synthesize ATP, but also contain endogenous ATPases that rapidly deplete the existing ATP. Luciferase reaction chemistry is used to quantify ATP, the limiting reactant, by generation of a luminescent signal that is proportional to viable cell number [32Maehara Y, Anai H, Tamada R, et al. The ATP assay is more sensitive than the succinate dehydrogenase inhibition test for predicting cell viability Eur J Cancer Clin Oncol 1987; 23: 273-76., 33Slater K. Cytotoxicity tests for high-throughput drug discovery Cur Opin Biotechnol 2001; 12: 70-4.].
All commercial ATP quantification reagents contain luciferase enzyme and luciferin substrate supplied with a lytic buffer. Cell lysis is necessary to liberate cellular ATP and is typically facilitated by detergents in the buffer. The most useful formulations for HTS applications also contain thermostable recombinant luciferase and ATPase inhibitors which allow for “glow-type” luminescence that produces a sustained signal with a half-life on the order of 5 hr [34Hall M, Gruber M, Hannah R, et al. Stabilization of firefly luciferase using directed evolution In: Roda A, Pazzagli M, Kricka L, Eds. Bioluminescence and Chemiluminescence—Perspectives for the 21st Century. Chichester UK: John Wiley & Sons 1998; pp. 392-5., 35Riss T. Selecting cell-based assays for drug discovery screening Cell Notes 2005; 13: 16-21.]. Prolonged maintenance of the luminescent signal allows for batch processing of plates and obviates any inter-plate kinetic variability.
The principle advantages of an ATP assay include that it is currently the most rapid and sensitive HTS method available for assessing viability. For instance, as few as 10 cells can be detected in limiting dilution series of eukaryotic cells within 10 minutes (Fig. 2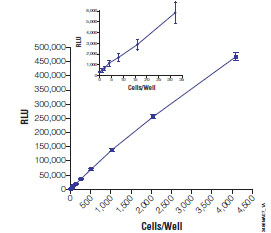 ). The assays also benefit from relatively high signal to background ratios, which enable routine miniaturization into 1536 well formats [36Severson W, Shindo N, Sosa M, et al. Development and validation of a high-throughput screen for inhibitors of SARS CoV and its application in screening of a 100,000-compound library J Biomol Screen 2007; 12: 33-40., 37Borawski J, Lindeman A, Buxton F, et al. Optimization procedure for small interfering RNA transfection in a 384-well format J Biomol Screen 2007; 12: 546-9.]. Furthermore, luminescence signals are not encumbered by intrinsically fluorescent test compounds [38Fan F, Wood K. Bioluminescent assays for high-throughput screening Assay Drug Dev Technol 2007; 5: 127-36.].
). The assays also benefit from relatively high signal to background ratios, which enable routine miniaturization into 1536 well formats [36Severson W, Shindo N, Sosa M, et al. Development and validation of a high-throughput screen for inhibitors of SARS CoV and its application in screening of a 100,000-compound library J Biomol Screen 2007; 12: 33-40., 37Borawski J, Lindeman A, Buxton F, et al. Optimization procedure for small interfering RNA transfection in a 384-well format J Biomol Screen 2007; 12: 546-9.]. Furthermore, luminescence signals are not encumbered by intrinsically fluorescent test compounds [38Fan F, Wood K. Bioluminescent assays for high-throughput screening Assay Drug Dev Technol 2007; 5: 127-36.].

Despite an abundance of advantages for HTS implementation, ATP assays also carry liabilities. Because the ATP-dependent luciferase reaction rate is susceptible to temperature flux, equilibration to constant room temperature is required to minimize thermal gradients created by plate well position (edge-effects). Also, like other metabolic assay chemistries used to measure viability, ATP is (rarely) subject to positive or negative modulation by test reagents independent of loss of viability leading to misleading data [39Sanchez-Alcazar J, Ruiz-Cabello J, Hernandez-Munoz I, et al. Tumor necrosis factor-α increases ATP content in metabolically inhibited L929 cells preceding cell death J Biol Chem 1997; 272: 30167-77., 40Shchepina L, Pletjushhkina O, Averisyan A, et al. Oligomycin, inhibitor of the Fo part of H+-ATP synthase, suppresses the TNF-induced apoptosis Oncogene 2002; 21: 8149-57.]. Lastly, all commercial formulations of ATP-detection chemistries are susceptible to luciferase inhibition to various degrees by small molecule compounds. Recent evidence suggests that specific formulations may greatly minimize the extent of this inhibition [41Kashem M, Nelson R, Yingling J, et al. Three mechanistically distinct kinase assays compared: measurement of intrinsic ATPase activity identified the most comprehensive set of ITK inhibitors J Biomol Screen 2007; 12: 70-83.].
CYTOTOXICITY ASSAYS
Cytotoxicity assay chemistries are formulated to detect loss of membrane integrity associated with cell death. The current most useful cytotoxicity biomarkers are constitutive, conserved and relatively stable, high-abundance enzymes “released” into the extra-cellular environment (culture medium) following loss of membrane integrity. Unlike viability assays whereby a decline in cell response is strongly inferred to be caused by cell-cycle arrest or death, detection of cytotoxic biomarker activities outside the cellular compartment after treatment is proof-positive for a compound’s cytotoxic effect. Unfortunately, activity-based surrogates of cell death have a finite enzymatic half-life, which can limit their utility during long compound/cell contact incubations (48-72 hr) [3Riss T, Moravec R. Use of multiple assay endpoints to investigate the effects of incubation time, dose of toxin, and plating density in cell-based cytotoxicity assays Assay Drug Dev Technol 2004; 2: 51-62., 27Niles A, Moravec R, Riss T. Update on in vitro cytotoxicity assays for drug development Expert Opin Drug Discov 2008; 3(6): 655-9.]. Enzymatic decay as a function of time in culture medium can lead to underestimation of the absolute level of cytotoxicity [27Niles A, Moravec R, Riss T. Update on in vitro cytotoxicity assays for drug development Expert Opin Drug Discov 2008; 3(6): 655-9.]. The quantitative HTS (qHTS) approach eliminates a majority of this error because the kinetics of cytotoxicity is intrinsically linked to dose [42Xia M, Huang R, Witt K, et al. Compound cytotoxicity profiling using quantitative high-throughput screening Environ Health Perspect 2008; 116(3): 284-91.].
LACTATE DEHYDROGENASE (LDH)
LDH activity has a long history as a preferred marker for cell death with in vitro cell models [43Korzeniewski C, Callewaert D. An enzyme-release assay for natural cytotoxicity J Immunol Methods 1983; 64: 211-4., 44Decker T, Lohmann-Matthes M. A quick and simple method for the quantification of lactate dehydrogenase release in measurements of cellular cytotoxicity and tumor necrosis factor (TNF) activity J Immunol Methods 1988; 115: 61-9.]. In HTS applications, the activity of LDH is indirectly measured by an enzymatically-coupled reagent chemistry comprised of lactate, NAD+, diaphorase and an appropriate redox dye such as a tetrazolium compound or resazurin. Although the result of LDH activity can be measured by a change in absorbance, fluorescence detection is favored because of greater sensitivity and the potential to miniaturize to high-density plates.
The implementation of LDH assays for cytotoxicity testing in HTS has been historically limited because most commercial assay protocols require a cell-free sample transferred into a separate assay plate. Reagents that are compatible with viable cells are available to detect release of LDH, thus enabling a single addition homogeneous assay format [45Riss T, Moravec R. Introducing the CytoTox-ONE homogeneous membrane integrity assay Cell Notes 2002; 4: 6-9.]. Now LDH release can be routinely measured in HTS formats after a short incubation (10-20 minutes) using a fluorometer equipped with a 560nm excitation source and a 590nm collection filter (Fig. 3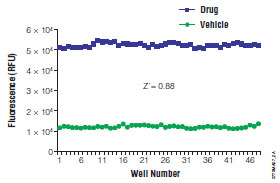 ) [46Bakshi P, Liao Y-F, Gao J, et al. A high-throughput screen to identify inhibitors of amyloid ß-protein precursor processing J Biomol Screen 2005; 10: 1-12.].
) [46Bakshi P, Liao Y-F, Gao J, et al. A high-throughput screen to identify inhibitors of amyloid ß-protein precursor processing J Biomol Screen 2005; 10: 1-12.].
 |
Fig. (3) LDH Assay Z’-factor Analysis. CytoTox-ONE® Assay reagent (Promega Corporation) was added to HepG2 cells treated with 3.125µM staurosporine. Medium served as vehicle control. |
LDH assay detection is a robust and cost-effective means of assessing cytotoxicity, but is susceptible to background signal from serum sources of LDH found in supplemented growth media [47Pei J, Ficht T. Brucella abortus rough mutants are cytopathic for macrophages in culture Infect Immun 2004; 72: 440-50., 48Hasinoff B, Schroeder P, Patel D. The metabolites of the cardioprotective drug dexrazoxane do not protect myocytes from doxorubicin-induced cytotoxicity Mol Pharmacol 2003; 64: 67.]. Exogenous sources of LDH greatly elevate background values and negatively impact the overall signal window. This experimental detriment can be largely mitigated by careful pre-screening of serum sources for cell culture prior to LDH assay. LDH assays are also subject to assay artifacts by compounds that inhibit the LDH activity and by standard fluorescence and color quenching interferences [49Anderson B, Noble C. In vitro inhibition of lactate dehydrogenases by kepone J Agric Food Chem 1977; 25: 8-31.-51Lash L, Zalups R. Mercuric chloride-induced cytotoxicity and compensatory hypertrophy in rat kidney proximal tubular cells J Pharmacol Exp Ther 1992; 261: 819-29.].
PROTEASE(S)
A specific proteolytic profile associated with cell death has been recently identified and can be measured in a manner similar to LDH assays [26Niles A, Moravec R, Hesselbeth P, et al. A homogeneous assay to measure live and dead cells in the same sample by detecting different protease markers Anal Biochem 2007; 366: 197-206.]. This method employs sequence-specific (Ala-Ala-Phe), fluorogenic or luminogenic peptide substrates delivered directly into assay wells via a buffered vehicle. The proteolytic activities present in viable cells are not detected by this method because normally compartmentalized proteases are not in contact with the impermeant substrates. However, protease(s) released into the culture medium as a result of cell death cleave the substrates releasing a detectible reporting molecule. The fluorescent format of the assay utilizes the highly-efficient rhodamine 110 (R110) fluorophore which is measured in as short as 30 minutes by a fluorometer with 485nm excitation and 520-30nm emission filters [52Liu J, Bhalgat M, Zhang C, et al. Fluorescent molecular probes V: a sensitive caspase-3 substrate for fluorometric assays Bioorg Med Chem Lett 1999; 9: 3231-6., 53Leytus S, Melhado L, Mangel W. Rhodamine-based compounds as fluorogenic substrates for serine proteinases Biochem J 1983; 209: 299-307.]. The luminescent format employs an engineered luciferase formulation to detect the luciferin product of cleavage thereby generating a glow-type signal which peaks at 15-20 minutes, stably persists for 3-4 hr, and is measured by standard luminometry.
Dead cell protease detection is advantageous in HTS and secondary screening applications because of improved sensitivity, linearity with respect to different degrees of cytotoxicity, reduction in background and artifactual hits, and choice in detection format (Fig. 4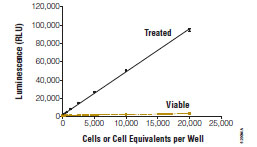 ). These attributes allow the technology to be applied in 1536 well formats as easily as in lower density plates [54Cho M-H, Niles A, Huang R, et al. A bioluminescent cytotoxicity assay for assessment of membrane integrity using a proteolytic biomarker Toxicol In Vitro 2008; 22: 1099-6.]. The dead cell protease detection method is subject to enzymatic activity decay due to the kinetics of cytotoxicity, biomarker inhibition, and standard interferences associated with the detection molecules.
). These attributes allow the technology to be applied in 1536 well formats as easily as in lower density plates [54Cho M-H, Niles A, Huang R, et al. A bioluminescent cytotoxicity assay for assessment of membrane integrity using a proteolytic biomarker Toxicol In Vitro 2008; 22: 1099-6.]. The dead cell protease detection method is subject to enzymatic activity decay due to the kinetics of cytotoxicity, biomarker inhibition, and standard interferences associated with the detection molecules.
 |
Fig. (4) Protease Assay Sensitivity and Linearity. CytoTox-Glo™ (Promega Corporation) was added to serial dilutions of viable and non-viable Jurkat cells and luminescence measured after 30 minutes. |
MULTIPLEXED VIABILITY AND CYTOTOXICITY ASSAYS
The single parameter assays described above offer convenient and predictive methods to view the cytotoxic landscape of chemical libraries. When certain combinations of the assays are added as same-well multiplexes however, they provide not only complementary but additive benefit [55Hallis T, Kopp A, Gibson J, et al. An improved β-lactamase reporter assay: multiplexing with a cytotoxicity readout for enhanced accuracy of hit identification J Biomol Screen 2007; 12: 635-44.]. Although the concept of multiplexed screening is well appreciated for image-based technologies, it is emerging as a more viable option for plate-based, homogeneous assays because implementation is simple, accessible, and cost effective [56Gerets HJ, Hanon E, Cornet M, et al. Selection of cytotoxicity markers for the screening of new chemical entities in a pharmaceutical context: a preliminary study using a multiplexing approach Toxicol In Vitro 2009; 23: 319-2.].
Reagent compatibility is a major consideration in same-well multiplexing. There is an absolute requirement that individual signal readouts are distinct and can be parsed for meaningful data set analysis. Also, reagent formulations must not compromise functional aspects of the chemistries. For instance, lytic assays for ATP measurement or caspase activation must be conducted in a sequence-specific order after first measuring experimental cytotoxicity.
Several informative combinations of viability and cytotoxicity chemistries are currently employed in HTS formats to mitigate false determinations. These multiplexes address systematic error caused either by disparate cell numbers delivered to assay wells or random compound interferences with reporter molecules [55Hallis T, Kopp A, Gibson J, et al. An improved β-lactamase reporter assay: multiplexing with a cytotoxicity readout for enhanced accuracy of hit identification J Biomol Screen 2007; 12: 635-44.]. Except in cases of cell-cycle arrest prior to loss of membrane integrity, or long term exposure models, measures of viability and cytotoxicity are inversely proportional (Fig. 5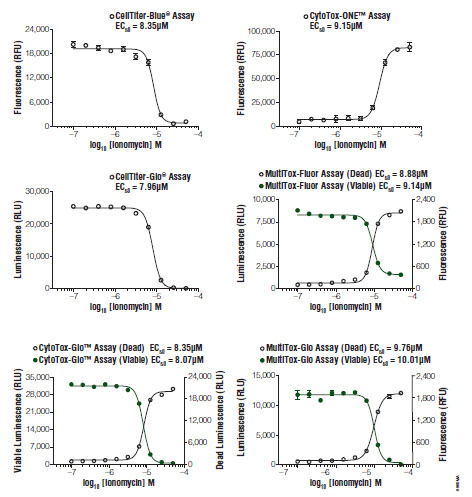 ). Therefore, non-conforming data sets can easily “flag” problem compounds for further analysis by orthogonal methods [57Niles A, Scurria M, Bernad L, et al. Measure relative numbers of live and dead cells and normalize assay data to cell number Cell Notes 2007; 18: 15-20.].
). Therefore, non-conforming data sets can easily “flag” problem compounds for further analysis by orthogonal methods [57Niles A, Scurria M, Bernad L, et al. Measure relative numbers of live and dead cells and normalize assay data to cell number Cell Notes 2007; 18: 15-20.].

FLUORESCENT/FLUORESCENT ASSAYS
Several classic examples of fluorescent viability and cytotoxicity assays are known [58Papadopoulous N, Dedoussis G, Spanakos G, et al. An improved fluorescence assay for the determination of lymphocyte-mediated cytotoxicity using flow cytometry J Immunol Methods 1994; 177: 101-1.]. These methods rely on an impermeant DNA intercalating dye which labels cells with damaged membranes and a permeable dye that is sequestered inside viable cells after esterase cleavage of a labeling group. Variations of these traditional chemistries are rarely employed where total fluorescence from assay wells is measured, because of poor sensitivity from simple stoichiometric dye binding events in the non-viable cell population and requisite washes required for removal of unbound dye and serum-derived esterases. Stable, non-viable cell esterase activity is particularly problematic for assay utility as it greatly impacts assay background values and constricts signal windows.
A new, more amenable method for HTS environments combines the protease substrates for viability and cytotoxicity allowing for simultaneous measurement of viable and non-viable cell populations (Fig. 6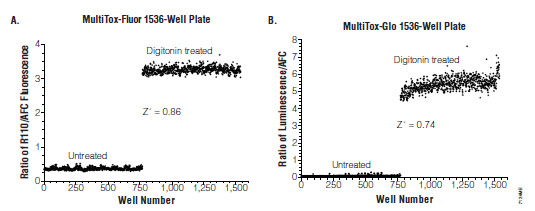 ). This method delivers broad linear responses (0-50,000 cells/well) and can measure as little as a 2% change in viability in both the AFC and R110 channels [59Niles A, Moravec R, Scurria M, et al. MultiTox-fluor multiplex cytotoxicity assay technology Cell Notes 2006; 15: 11-5.]. Furthermore, the reagent can be concentrated and delivered into assay wells in a substantially reduced volume to accommodate a third spectrally distinct reagent.
). This method delivers broad linear responses (0-50,000 cells/well) and can measure as little as a 2% change in viability in both the AFC and R110 channels [59Niles A, Moravec R, Scurria M, et al. MultiTox-fluor multiplex cytotoxicity assay technology Cell Notes 2006; 15: 11-5.]. Furthermore, the reagent can be concentrated and delivered into assay wells in a substantially reduced volume to accommodate a third spectrally distinct reagent.

Triplexes created by uniting the fluorescent viability and cytotoxicity multiplex with luminescent chemistries are advantageous for several reasons. First, it is possible to conduct immediate confirmation testing of apparent cytotoxic compounds in the same well by leveraging the statistical power of orthogonal detection of viability (viable cell aminopeptidase, dead cell protease and ATP biomarkers). The economy of this consolidated testing is noteworthy in terms of compound usage, culture set up, and operator man-hours. Second, the mechanism of cell death can often be elucidated when combined with caspase detection reagents, allowing for partial characterization of a compound’s potential. It should be noted however, that caspase activity can be transient and immeasurable in long-term experiments due to the kinetics of apoptosis and caspase biomarker decay. Therefore, caspase-negative data sets should be rescreened at multiple compound exposure periods. This type of characterization of pathway dependent cytotoxicity is useful for continued annotation of a library, because compounds causing necrotic profiles at micromolar concentrations are rarely therapeutically useful and may offer unacceptable cytotoxic risk. And third, viability and cytotoxicity can be used to normalize luminescent data from other assays (Fig. 7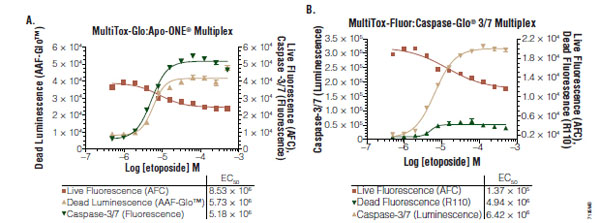 ). Single parameter genetic reporter responses are susceptible to experimental variation due to differential transfection efficiencies, cell number delivered to the well (clumping) or to cytotoxicity caused by the test compound [55Hallis T, Kopp A, Gibson J, et al. An improved β-lactamase reporter assay: multiplexing with a cytotoxicity readout for enhanced accuracy of hit identification J Biomol Screen 2007; 12: 635-44.]. Viability and cytotoxicity data can therefore help reconcile aberrant data points or explain decreases in genetic reporter signals due to cell death.
). Single parameter genetic reporter responses are susceptible to experimental variation due to differential transfection efficiencies, cell number delivered to the well (clumping) or to cytotoxicity caused by the test compound [55Hallis T, Kopp A, Gibson J, et al. An improved β-lactamase reporter assay: multiplexing with a cytotoxicity readout for enhanced accuracy of hit identification J Biomol Screen 2007; 12: 635-44.]. Viability and cytotoxicity data can therefore help reconcile aberrant data points or explain decreases in genetic reporter signals due to cell death.

FLUORESCENT/LUMINESCENT ASSAYS
Mixed detection combinations of the protease biomarkers are also available that offer benefit in screening applications. In this method, viability is measured first by the addition of the GF-AFC reagent followed by the addition of the luminogenic dead cell reagent chemistry. The use of disparate detection platform methodologies is known to greatly reduce the statistical potential for assay interferences associated with reporter molecules [60Grant S, Sklar J, Cummings R. Development of novel assays for proteolytic enzymes using rhodamine-based fluorogenic substrates J Biomol Screen 2002; 7: 531-40.]. Therefore this combination of chemistries is favored if a library is uncharacterized with regard to the relative abundance of luminogenic inhibitors or which may cause fluorescence interference. Unlike the fluorescence/fluorescence multiplex, fewer convenient triplex options exist. For example, the chemistry can be configured with a fluorogenic caspase activation chemistry (R110 fluorophore) to deliver data relating to possible induction of apoptosis (Fig. 7A) [61Worzella T, Busch M, Niles A. High throughput automation of multiplexed cell viability assays for viability and cytotoxicity Cell Notes 2008; 20: 26-9., 62Niles A, Moravec R, Riss T. Multiplex caspase activity and cytotoxicity assays Methods Mol Biol 2008; 414: 151-62.]. Again, negative caspase induction results should be verified by rescreening to verify necrotic response profiles.
LUMINESCENT/LUMINESCENT ASSAYS
Fully-luminescent formats are highly desirable for HTS environments because of the aforementioned advantages over fluorescent methods. Another remarkable feature of luminogenic assays is the persistent and stable glow-type signal produced after reagent addition (Fig. 8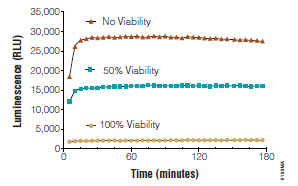 ). This attribute can be harnessed to produce both viability and cytotoxicity values from the same well. In this method, the non-lytic, luminogenic cytotoxicity assay chemistry is first delivered to the assay well and cytotoxicity measured. Next, a lytic agent is added to assay wells to artificially release the remaining dead cell protease from viable cells. The net signal derived after complete lysis reflects the contribution of all the cells in the assay well. Because the luminescent signal is additive and proportional, the viable cell population can be indirectly determined by subtracting the cytotoxic values from the total values. This method is particularly useful for determining cytotoxicity in long incubation protocols where dead cell biomarker activity may not fully reflect actual cytotoxicity. One caveat of this method is that both viability and cytotoxicity measurements utilize the same biomarker and are subject to the same interferences.
). This attribute can be harnessed to produce both viability and cytotoxicity values from the same well. In this method, the non-lytic, luminogenic cytotoxicity assay chemistry is first delivered to the assay well and cytotoxicity measured. Next, a lytic agent is added to assay wells to artificially release the remaining dead cell protease from viable cells. The net signal derived after complete lysis reflects the contribution of all the cells in the assay well. Because the luminescent signal is additive and proportional, the viable cell population can be indirectly determined by subtracting the cytotoxic values from the total values. This method is particularly useful for determining cytotoxicity in long incubation protocols where dead cell biomarker activity may not fully reflect actual cytotoxicity. One caveat of this method is that both viability and cytotoxicity measurements utilize the same biomarker and are subject to the same interferences.

SUMMARY AND CONCLUSIONS
In vitro cytotoxicity testing technologies for HTS continue their refinement on many fronts. High Content Screening (HCS) and label-free methods have gained some degree of popular following recently, but require expensive instrumentation, complicated analysis software packages and offer relatively low throughput. With burgeoning costs associated with drug discovery efforts, straight-forward, validated, homogeneous assay chemistries remain the choice as a simple and cost-effective means of assessing cytotoxic burden.
Depending upon the goals of the screen, either viability or true cytotoxicity assay chemistries can be employed. Cytotoxicity assays based on membrane integrity changes are positive-readout assays most typically indicated for shorter-term exposure models (48 hr or less). These assays may not always accurately estimate the absolute degree of early or late stage cytotoxicity due to the kinetics of biomarker emergence or degradation. Viability assays measure the level of biomarker activity inversely correlated with cytotoxicity and therefore may be used at any endpoint during a compound/cell incubation period.
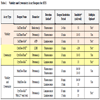
Each biomarker of viability and cytotoxicity has advantages and disadvantages. The choice of which surrogate of health or death to use is greatly influenced by the available detection format and extent of assay time required to reach a result (Table 2). Fluorescent formats require short to lengthy incubations with the sample whereas luminescent formats tend to deliver maximal signal windows in a rapid but prolonged fashion. Luminescent formats offer additional utility over fluorescent methods with regard to detection sensitivity, but are also typically more costly to use on a per well basis.
All assay chemistries are subject to experimental error or interferences which can lead to false interpretation of data sets. Multiplexed combinations of compatible assays offer a convenient and cost-effective manner to address variation (by response normalization), flag non-conforming orthogonal data points and increase per well content. Although greatly informative, special attention should be paid to experimental design and the interpretation of multiplexed data sets due to the transient or kinetic nature of mechanistic biomarkers.
ACKNOWLEDGEMENT
The authors would like to thank Michele Arduengo, Ph.D. for preparing and providing the figures for this work.