- Home
- About Journals
-
Information for Authors/ReviewersEditorial Policies
Publication Fee
Publication Cycle - Process Flowchart
Online Manuscript Submission and Tracking System
Publishing Ethics and Rectitude
Authorship
Author Benefits
Reviewer Guidelines
Guest Editor Guidelines
Peer Review Workflow
Quick Track Option
Copyediting Services
Bentham Open Membership
Bentham Open Advisory Board
Archiving Policies
Fabricating and Stating False Information
Post Publication Discussions and Corrections
Editorial Management
Advertise With Us
Funding Agencies
Rate List
Kudos
General FAQs
Special Fee Waivers and Discounts
- Contact
- Help
- About Us
- Search


The Open Anatomy Journal
(Discontinued)
ISSN: 1877-6094 ― Volume 6, 2014
En Bloc Removal of the Mandible, the Masticatory Muscles and the Mandibular Nerve in the Chimpanzee (Pan troglodytes, Blumenbach 1799), with a Review and Critique of the Methods Used to Expose the Trigeminal Musculature in Derived Primates
M Ashraf Aziz1, Kerrie Lashley1, Cecile E Skinner2, Rui Diogo*, 3
Abstract
The detailed protocol of the en bloc removal of the mandible, the muscles supplied by the mandibular nerve tree and the maxillary artery is described in the chimpanzee (Pan troglodytes). This new method allows integrated observations of the masticatory muscles, their ontogenetic associates and the specific branches of the mandibular nerve that supply them. The topographic relationships of the muscles, including their subparts, are seen in light of the specific nerve branches that supply and interconnect them. Previous methods that examine parts of the trigeminal musculature and the mandibular nerve in a fragmented manner are described and critiqued. The present method allows us to arrive at a sounder classification of the trigeminal muscles based on the integrated observation of their nerve supply; this approach was first proposed by Toldt and more recently used by Tomo. The anatomical, functional, clinical, ontogenetic and phylogenetic aspects of the new approach are discussed.
Article Information
Identifiers and Pagination:
Year: 2011Volume: 3
First Page: 21
Last Page: 33
Publisher Id: TOANATJ-3-21
DOI: 10.2174/1877609401103010021
Article History:
Received Date: 10/6/2011Revision Received Date: 1/7/2011
Acceptance Date: 4/7/2011
Electronic publication date: 1/8/2011
Collection year: 2011
open-access license: This is an open access article licensed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted, non-commercial use, distribution and reproduction in any medium, provided the work is properly cited.
* Address correspondence to this author at the Center for the Advanced Study of Hominid Paleobiology, Department of Anthropology, George Washington University, 2110 G Street, N.W., Washington, DC 20052, USA; Tel: 2026510439; Fax: 2029946097; E-mails: rui_diogo@hotmail.com, ruidiogo@gwmail.gwu.edu
| Open Peer Review Details | |||
|---|---|---|---|
| Manuscript submitted on 10-6-2011 |
Original Manuscript | En Bloc Removal of the Mandible, the Masticatory Muscles and the Mandibular Nerve in the Chimpanzee (Pan troglodytes, Blumenbach 1799), with a Review and Critique of the Methods Used to Expose the Trigeminal Musculature in Derived Primates | |
INTRODUCTION
Since the beginning of morphological investigations on the head myology of the chimpanzee, its muscles of facial expression have received the greatest scrutiny [1-10]. More recently, Burrows et al. [11] developed the novel “face mask” dissection method to elucidate hitherto unrecognized morphological complexity of the facial muscles in the chimpanzee. The “face mask” dissection has elucidated the functional morphology of the facial muscles, leading to fresher insights regarding this important group of cranial muscles of communication.
Curiously, another equally prominent group of head muscles - those that control the actions of the temporomandibular joint and their myological relations - have yet to be subjected to equally detailed, comprehensive study. Sonntag [4] provides an early summary of what was then known of these muscles. He wrote a sketchy description of the masseter and the temporalis; with reference to the pterygoid muscles he writes that “they are disposed as in man, and their relations to the vessels and nerves in the pterygoid space are similar”. His book includes a flat, two-dimensional (i.e., devoid of any artistic techniques suggesting depth) diagram summarizing a number of superficial yet salient aspects of “the pterygoid…and palatine..regions” of the Hominodea. It is not clear if the schematic diagram actually refers to the structures as seen in a dissected chimpanzee or represent a composite of those structures as seen in humans and/or other hominoids.
Yoshikawa and Suzuki [12] and Gaspard et al. [13] have reported on the detailed lamination and organization of the masseter in the chimpanzee. Göllner [14] and Swindler and Wood [15] have provided a somewhat detailed and comprehensive description of the masticatory muscles that are innervated by the trigeminal nerve (Cranial Nerve V) in adult and neonatal chimpanzees. However, these investigations do not include other, “non-masticatory” head muscles that are also supplied by the mandibular branch of the trigeminal nerve. Despite Göllner’s [14] efforts to provide a detailed description of the masticatory muscles and the branches of the mandibular nerve (the only branch of the trigeminal nerve that actually supplies these muscles), he did not map them in the way that Schwartz and Huelke [16], Tomo [17], Tomo et al. [18, 19] and Aziz et al. [20] have done in the rhesus monkey, the dog, and in humans, respectively.
Traditionally, the masticatory muscles have been described as distinct muscles enclosed in their fascial compartments. Furthermore, the descriptions include their positional relationships to each other, and the precise attachments of their tendons. However, the painstaking comparative studies using such an approach have led to unsatisfactory results [21-27]. Of particular concern have been those muscle bundles that have been found between the masseter and the temporalis, on one hand, and between the temporalis and the lateral pterygoid, on the other [28-30]. Furthermore, there is a lack of agreement regarding the partitions of each specific masticatory muscle (e.g., the masseter, medial pterygoid, etc.) [21-28].
Toldt [31] was among the first investigators to study the comparative morphology of the masticatory muscles based on their nerve supply. This approach has been further developed by other authors [32-43]. Lubosch [37] referred to the muscles supplied by the mandibular branch of the trigeminal nerve as “trigeminusmuskulatur”. His investigations involved fish, amphibians and reptiles. Tomo extended Toldt’s classification of the masticatory muscles based on their specific nerve supply from the mandibular nerve to mammals, in particular the dog [17-19]. Göllner [14] used this approach in his description of selected craniomandibular muscles including the masticatory muscles of the chimpanzee and the gorilla. These investigations have provided a strong justification for the nerve supply-based classification of the masticatory muscles (including isolated muscles or muscle bundles that accompany the commonly named muscles of mastication) [44-47] and their subparts [29, 30, 48, 49]. The other advantage of such an approach to the classification of the masticatory muscles is that it elucidates their ontogenetic and phylogenetic relationships [17-19]. Clearly, this is the biologically rational method for classifying muscles because it is grounded in evolutionary studies.
However, all the above-mentioned studies relate to the masticatory muscles and their immediate functional associates (i.e. the anterior digastric and the mylohyoid) only. They have not been extended to the entire myological field of the mandibular nerve tree which, in mammals, in addition to the muscles of mastication and their functional associates, also includes a muscle of the soft palate (tensor veli palatini) and an auditory muscle (tensor tympani) [10, 35, 36, 42, 43, 50, 51]. The “trigeminal musculature” – or mandibular musculature sensu Diogo et al. [10] - comprises muscles of branchiomeric origin; proximally, these muscles differentiate from a common mesodermal plate associated with the embryonic mandibular arch (1st branchial arch). In humans, the ultimate source of these so-called “jaw closing” muscles is somitomere 4 which originates from the cephalic portion of the paraxial mesoderm [52]. Anomalous morphogenesis of the mandibular arch plate in the period between the 4th and 6th weeks can lead to anomalies of all or most structures – bones, muscles, nerves, and vasculature – derived from or associated with the mesodermal plate of that arch [52-54]. Clearly, it is more meaningful to investigate the “trigeminal musculature” as a lineage of muscles which arise from a common source under the control of a set of functionally related genes [10].
A major impediment to the study of the “trigeminal musculature” as an integrated unit has been the difficulty in accessing its components as a whole. The masseter and the temporalis are the best studied parts of this unit because large sections of these muscles are easily dissectable due to their superficial location [24-27, 29, 30, 48, 49]. The pterygoids, the tensor veli palatini and especially the tensor tympani have posed challenges because they are enclosed in spaces bounded by bony plates which require a complicated series of osteotomies for full exposure [20, 50-60]. The piecemeal exposure of parts of the infratemporal fossa, the pterygoid fossa and the auditory tube have prevented investigators from viewing these muscles as distinct parts that can be arranged – individually or collectively – into an ontogenetic (and phylogenetic) sequence [17-19, 48, 49].
A major impediment to the detailed and comprehensive study of the “trigeminal musculature” is thus an issue of methodology. Even when these muscles have been dissected on a piecemeal basis, the methods – a few exceptions notwithstanding [55, 56] – of exposing the deep-seated “trigeminal muscles” and their neurovascular associates have not been described with sufficient clarity. Therefore, subsequent studies have faced difficulty in reproducing the earlier work with precision. Then, too, the methods which have been used so far only allow restricted parts of the masticatory or other neighboring spaces to be exposed; this prevents us from seeing these structures as integrated components derived from a common source. Another reason for the fragmentary description of the “trigeminal musculature” is the need to divide the head anatomy into regions that are associated with the dissection-based study and instruction [61-63]. Thus, the masticatory muscles, the muscles of the floor of the oral cavity, the muscles of the soft palate and the otic muscles are dissected and studied as isolated subregions. There is seldom an attempt to integrate the information and see them as a single developmental neuromuscular field.
The objective of our paper is to describe a method by which the entire mandible, the TMJ disc, the masticatory muscles, their functional associates, the tensor veli palatini, the entire mandibular nerve tree, and the proximal parts of the maxillary artery can be removed en bloc and studied as an interconnected set of functional components; within the “trigeminal muscles”, only the tensor tympani remains inaccessible using this method (although there are ways by which it too can be extracted along with its relations: we plan to do this is a future work). The advantages of this method and its rationale are discussed. Existing methods which have been used to investigate the muscles supplied by the trigeminal nerve in humans and nonhuman primates are described and critically evaluated. To our knowledge, this is the first time that the mandible, its muscles and their nerves (including the accompanying vessels) has been removed en bloc and then studied in detail in the chimpanzee.
MATERIALS AND METHODS
The head of a post-autopsied chimpanzee (Pan troglodytes) adult male was acquired from the Yerkes Regional Primate Research Center (Emory University, Atlanta, GA). It was designated Specimen HUC 102; it is a part of a collection currently being used to dissect and map the infratemporal fossa of the chimpanzee in greater detail. HUC 102 had already undergone craniotomy to remove the brain for neuro-anatomical investigations; the orbits, too, had been emptied of their contents (Fig. 1A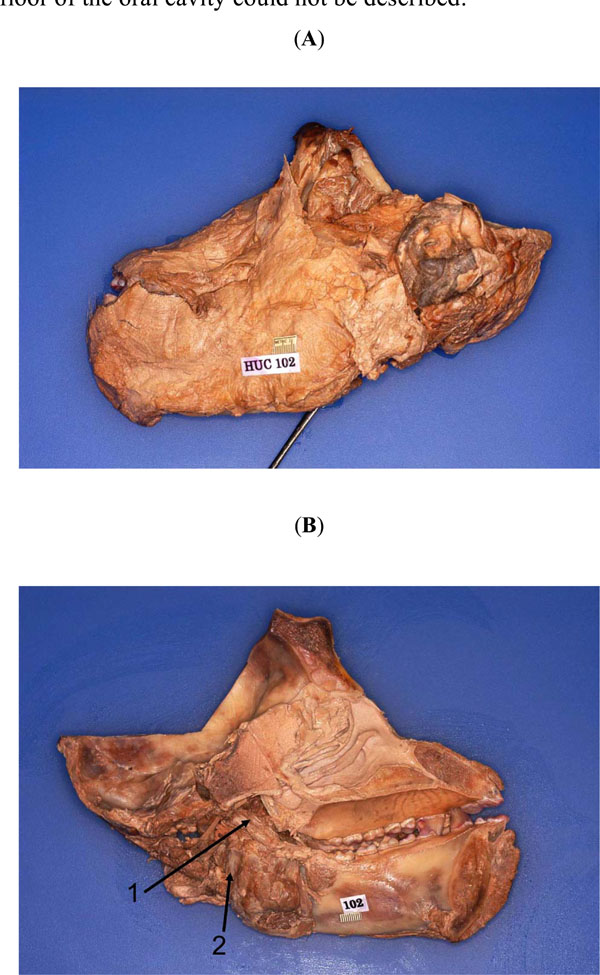 , B). The tongue, along with the surrounding oral cavity structures located at the floor (muscles, nerves, glands, etc.) had also been excised during the autopsy. However, the parotid gland and its duct, as well as its other contents were intact; the facial muscles and their nerves were also found to be undisturbed; only the muscles supplied by the trigeminal nerve in the floor of the oral cavity could not be described.
, B). The tongue, along with the surrounding oral cavity structures located at the floor (muscles, nerves, glands, etc.) had also been excised during the autopsy. However, the parotid gland and its duct, as well as its other contents were intact; the facial muscles and their nerves were also found to be undisturbed; only the muscles supplied by the trigeminal nerve in the floor of the oral cavity could not be described.

The other masticatory muscles and their neurovascular associates were intact; only the upper portions of the temporalis had been cut during the craniotomy. The lower parts of the temporalis, including its insertion tendon, were intact, bilaterally. Thus, HUC 102 presented an excellent opportunity to attempt a complete removal of the mandible, along with its masticatory muscles, and their neurovascular associates.
Prior to dissection, the head was bisected using an electrical bandsaw. This facilitated the exposure of the medial pterygoid and the tensor veli palatini and their neurovascular supplies. The bisection also opened up the plane from which the foramen ovale could be approached later – using osteotomy - to mobilize the mandibular nerve and the middle meningeal artery.
In addition to the traditional dissecting instruments commonly used in the gross anatomy laboratory, the following tools were essential to achieve optimal results: stryker saw; fluorescent magnifier lamp (1.75x); Hu-Friedy hard wire cutter (or any similar bone cutter); Blumenthal 30 rongeur, and - important - the periosteal elevator (Hu-Friedy #9 molt). At critical phases the dissection process was halted to permit photography, hand drawings, and note taking. The same dissection was performed on both sides.
After the removal of the mandible, the masticatory muscles and their nerves and vessels, further dissection was conducted under the fluorescent magnifying lamp using several microsurgical instruments, such as microscalpels, microscissors, the pin probe, and dissection pins. The magnifying lamp is essential to facilitate the unequivocal exposure of the finer nerves and vessels that are located deep to the lateral pterygoid heads. It is in this area that the mandibular nerve descends through the foramen ovale into the roof of the infratemporal fossa where it immediately bifurcates giving rise to its “anterior” and “posterior” divisions or trunks [64]. An important branch of the “anterior” division known as the buccal nerve (also known as the long buccal nerve) travels in the cleft between the superior and inferior heads of the lateral pterygoid. Additionally, the maxillary artery and its branches and the pterygoid venous plexus and its tributaries are very closely associated with the same muscle.
To complement our observations of the soft-tissues, we carefully examined an actual bony skull of another chimpanzee from our collection in order to visualize the infratemporal fossa (and its bony parts) and locate it with reference to neighboring spaces, especially the middle cranial fossa which housed the trigeminal nerve (CN V, and its divisions) and its sensory (Gasserian) ganglion. Since the anatomy of specific areas of the skull is critical for our protocol, the following areas were studied in detail: 1) The Middle Cranial Fossa: the superior orbital fissure; foramen rotundum; foramen ovale; Meckel’s cave or fossa; carotid canal; (foramen spinosum); grooves of the middle meningeal artery; and petro-sphenoid suture (Fig. 2A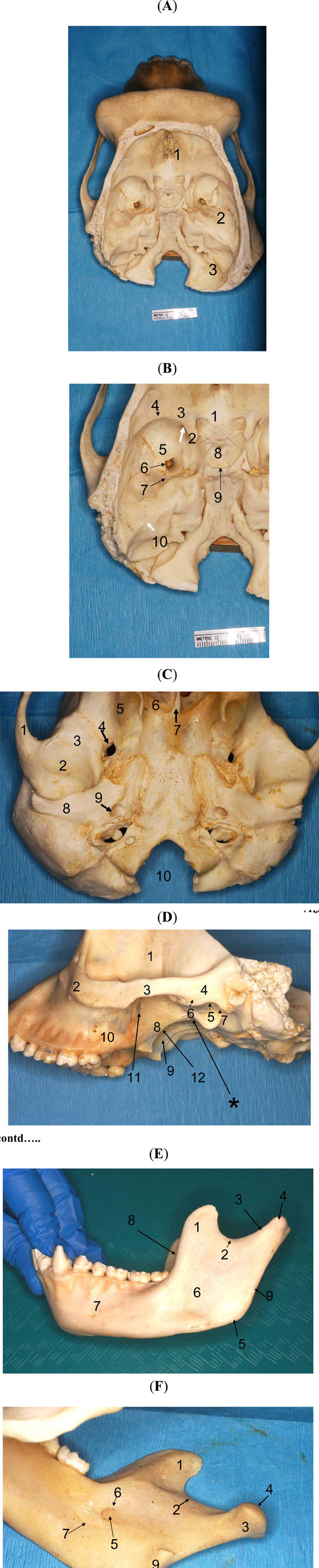 , B); 2) The Skull Base (Basicranium; External Surface) below the Infratemporal Crest of the Sphenoid: fossa (or impression) of the superior lateral pterygoid; external pterygoid plate; pterygoid hamulus; pterygoid fossa; foramen ovale; auditory tube; sphenoid “spine” (actually tuberosity); mandibular fossa; articular tubercle; postglenoid tubercle; squamotympanic fissure; petrotympanic fissure; and the scaphoid process (Fig. 2C); 3) The Lateral Aspect of the Skull : maxilla (posterior aspect); maxillary tuberosity; palatine (pyramidal process); inferior orbital fissure; pterygomaxillary fissure; lateral pterygoid plate; medial pterygoid plate; pterygoid hamulus; pterygoid fossa; foramen ovale; medial buttress of the mandibular fossa (Fig. 2D); 4) The Mandibular Ramus: coronoid process; mandibular (sigmoid) notch; condyle (head; neck); masseteric depression; medial pterygoid fossa; mylohyoid groove; mandibular foramen/canal; lingula; depression of the temporalis; and the angle of mandible (Fig. 2E, F).
, B); 2) The Skull Base (Basicranium; External Surface) below the Infratemporal Crest of the Sphenoid: fossa (or impression) of the superior lateral pterygoid; external pterygoid plate; pterygoid hamulus; pterygoid fossa; foramen ovale; auditory tube; sphenoid “spine” (actually tuberosity); mandibular fossa; articular tubercle; postglenoid tubercle; squamotympanic fissure; petrotympanic fissure; and the scaphoid process (Fig. 2C); 3) The Lateral Aspect of the Skull : maxilla (posterior aspect); maxillary tuberosity; palatine (pyramidal process); inferior orbital fissure; pterygomaxillary fissure; lateral pterygoid plate; medial pterygoid plate; pterygoid hamulus; pterygoid fossa; foramen ovale; medial buttress of the mandibular fossa (Fig. 2D); 4) The Mandibular Ramus: coronoid process; mandibular (sigmoid) notch; condyle (head; neck); masseteric depression; medial pterygoid fossa; mylohyoid groove; mandibular foramen/canal; lingula; depression of the temporalis; and the angle of mandible (Fig. 2E, F).

RESULTS
The en bloc removal of the mandible, the masticatory muscles, the mandibular nerve (and its branches), the maxillary artery (and its branches), and the maxillary vein (and its tributaries) was achieved in the following sequential steps:
Step 1: The zygomaticus major and minor, the platysma and the associated facial muscles and their fasciae were mobilized by dissection and reflected in the rostral direction. The outer surfaces of the zygoma and the zygomatic arch were exposed by removing all the fasciae and the periosteal membrane; cleaning was extended all the way posteriorly as close to the external auditory meatus as possible (Fig. 3 ). The parotid duct and the buccal nerves (CN VII) were exposed as they coursed below the zygomatic arch. These were cut close to their origin from the parotid gland; they, too, were reflected in the same direction as the facial muscles and fasciae described above. The temporal fascia was detached from the temporalis (the superior part of which had mostly been removed during the craniotomy), and dissected down to its attachment along the zygomatic bone and arch. This fascia was cut and removed.
). The parotid duct and the buccal nerves (CN VII) were exposed as they coursed below the zygomatic arch. These were cut close to their origin from the parotid gland; they, too, were reflected in the same direction as the facial muscles and fasciae described above. The temporal fascia was detached from the temporalis (the superior part of which had mostly been removed during the craniotomy), and dissected down to its attachment along the zygomatic bone and arch. This fascia was cut and removed.

Step 2: The borders of the parotid gland were more clearly defined by the removal of the binding connective tissue along the borders. The gland was pried away from the masseter and lifted up along its anterior border and reflected rostro-dorsally towards the ear. It was lifted clear of its attachment along the posterior margin of the mandibular ramus. Thus, the retromandibular fossa/space was exposed. The glandular tissue in the area just below the temporomandibular joint (TMJ) was teased up in order to expose the auriculotemporal nerve, the external carotid artery (and its branches: the maxillary and the superficial temporal arteries), and the retromandibular vein (and its tributaries: the maxillary vein and the superficial temporal vein).
Step 3: The auricle of the ear was excised close to its attachment to the outer, cartilaginous margins of the external auditory meatus. Thus, the TMJ’s external surface, with its capsule, and ligament were now revealed. The whole area was cleaned in order to better delineate these structures; cleaning was extended to the posterior aspect of the joint. The membranous “stylomandibular” ligament was found inserting on the posterior aspect of the mandibular ramus, just above the mandibular angle.
Step 4: The external surface of the masseter was cleaned and its borders defined. Using the spatulate end of the periosteal elevator, the muscle was detached from its insertion on the outer aspect of the mandibular ramus. This detachment was continued towards the zygomatic arch. The muscle was lifted, and by gentle teasing in the area close to the (over the mandibular notch) TMJ, the masseteric nerve, artery and vein were located and defined as they divided and arborized in the muscle. The zygomatic arch was now mobilized by the following two cuts using the stryker saw: an oblique cut of the zygomatic bone and a cut of the zygomatic process of the temporal bone just anterior to external auditory meatus (Fig. 3). The zygomatic arch was pulled laterally to expose the deep masseter and the “maxillomandibularis” muscle [27]; some of the fibers of the “maxillomandibularis” were attached close to the temporalis tendon. In the vicinity of the TMJ, the deep masseter and the temporalis appeared to be continuous.
Step 5: The temporalis was mobilized from the temporal fossa by using the spatulate end of the periosteal elevator. This process was extended all the way to the posterior margin of the frontal process of the zygomatic. Muscle detachment was continued as far inferiorly and caudally as possible. The anterior deep temporal artery and its branches could be seen after the fatty tissue (“corpus adiposum buccae”) covering the anterior margin of the zygomaticomandibularis [10, 27] was cleaned. The temporalis was pulled laterally to expose the anterior deep temporal artery as it coursed antero-superiorly towards the border of the zygomaticomandibularis. The lower part of the temporalis in the vicinity of the infratemporal crest of the greater wing of the sphenoid was teased to expose the deep temporal nerves which were accompanied by the branches of the posterior deep temporal artery.
Step 6: The hemi-section was now turned over to expose its medial aspect. The medial pterygoid’s superficial surface was cleaned and its boundaries delineated.
Step 7: The soft palate was cut and removed. This exposed the rostro-superior aspect of medial pterygoid. It also exposed the tensor veli palatini which was located in the pterygoid fossa. The medial pterygoid was detached from its insertion on the medial aspect of the mandibular ramus. The muscle was mobilized along its posterior border. As this process was extended rostro-superiorly, the mylohyoid nerve and vessels could be seen attached to the deep/external aspect of the medial pterygoid. This neurovascular bundle was pried away from the muscle and laid on to the mandibular ramus. The muscle detachment was extended antero-superiorly to reveal the inferior alveolar neurovascular bundle as it entered the mandibular canal through the mandibular foramen. The spheno- mandibular ligament was clearly visible in this area. Further muscle mobilization in the rostral direction exposed the lingual nerve as it coursed toward the floor of the oral cavity; it traversed along the medial margin of the mandibular ramus and its body. It was accompanied by a lingual branch of the inferior alveolar artery.
Step 8: The tensor veli palatini was pried away from the medial pterygoid to reveal the nerve to the medial pterygoid. A smaller nerve, located immediately to its rostro-superior aspect, was seen feeding into the tensor veli palatini.
Step 9: The tensor veli palatini and the medial pterygoid were now detached from the pterygoid fossa. The pointed end of the periosteal retractor is an excellent tool to achieve this. Still, at some points, where the muscles were very strongly attached to the bone, the microscalpel was used to detach the muscle. The external part of the medial pterygoid was reflected by cutting along the external surface of the lateral pterygoid plate. This cut was extended upward to mobilize the origin of the inferior head of the lateral pterygoid muscle.
Step 10: The TMJ capsule on the medial aspect of the joint was now cleaned, exposed and cut horizontally. The cut was no more than 4mm from the posterior margin. This limited cut prevented the excision of the main stump of the mandibular nerve as it emerged from the foramen ovale into the infratemporal fossa.
Step 11: The hemi-head was now turned to bring the internal floor of the basicranium (i.e. the view showing the cranial fossae) into full view. The dura mater was peeled away to expose the entire trigeminal nerve, its sensory ganglion, and its three divisions (ophthalmic, maxillary, and mandibular nerves). The entire nerve was cleaned and its borders defined. The nerve, its ganglion, and its divisions (ophthalmic, maxillary, and mandibular) were now peeled away from medial wall of the middle cranial fossa (greater wing of sphenoid). The ophthalmic and maxillary nerves were cut and freed in the immediate vicinity of their entry into the superior orbital fissure and the foramen rotundum, respectively.
Now, a wedge-cut pointing towards the foramen ovale (containing the mandibular division of the trigeminal nerve and other structures) was made from the surface using the stryker saw (Fig. 4 ). The anterior cut was made starting from the area close to the sphenoparietal eminence - the area close to the optic canal. The posterior cut began in the posterior cranial fossa and was continued through the petrous temporal bone towards the foramen ovale. In order to preserve the mandibular nerve in the foramen ovale; care was taken not to cut into it. Rather, the cuts reached close to the foramen in order to preserve the deep boundary of the opening. The trigeminal nerve was retracted laterally during the osteotomy to prevent damage to it. Now, using the pliers, the remaining bony wedge close to the foramen ovale was broken off. The rongeur was used to remove the remaining bone along the medial border of the foramen ovale; the area around the main stump of the mandibular nerve was cleaned to mobilize it along with vessels. The entire freed portion of the mandibular nerve (and the rest of the trigeminal nerve) was now pried out of the foramen ovale.
). The anterior cut was made starting from the area close to the sphenoparietal eminence - the area close to the optic canal. The posterior cut began in the posterior cranial fossa and was continued through the petrous temporal bone towards the foramen ovale. In order to preserve the mandibular nerve in the foramen ovale; care was taken not to cut into it. Rather, the cuts reached close to the foramen in order to preserve the deep boundary of the opening. The trigeminal nerve was retracted laterally during the osteotomy to prevent damage to it. Now, using the pliers, the remaining bony wedge close to the foramen ovale was broken off. The rongeur was used to remove the remaining bone along the medial border of the foramen ovale; the area around the main stump of the mandibular nerve was cleaned to mobilize it along with vessels. The entire freed portion of the mandibular nerve (and the rest of the trigeminal nerve) was now pried out of the foramen ovale.

Step 12: The spatulate end of the periosteal retractor was inserted into the superior cavity of the TMJ. The cavity was opened by pushing the retractor caudally. This revealed the attached anterior part of the TMJ capsule. It was very carefully cut and the retractor was pushed forward and downward to peel the attachment of the superior lateral pterygoid from the infratemporal part of the greater wing of the sphenoid. The nerves to the masseter, the posterior and middle deep temporal nerves, and the (long) buccal nerve were separated out along with the superior pterygoid head.
Step 13: A cut was made from the posterior edge of the oral cavity to divide the cheek in order to free the mandible. A careful inspection was made of all sides to free all structures that still articulated / connected the mandible to the cranium. Now, using measured force (and ensuring that the stump of the mandibular was clearly out of the foramen ovale), the mandible was pried away from the cranium (Fig. 5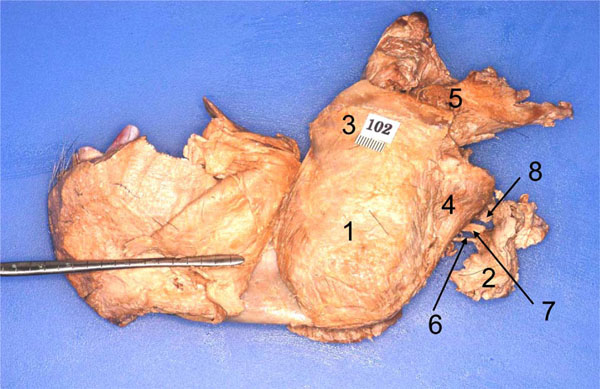 ).
).

Step 14: The entire mandible, its muscles, and neurovascular structures were cleaned by removing the covering fasciae. Following this the mandible was brought under the illuminated magnifier to extend the dissection to the finer structures of the entire complex (Fig. 6 ).
).
As explained above, the main goal of this paper is not to describe all the details of the masticatory muscles, the mandibular nerve and its branches, and the associated vasculature. Those details will be extensively described in a future publication. For the purposes of the present paper, we would just like to note that, in general, our observations of the soft masticatory structures in the chimpanzee (see, e.g. Fig. 6) are in accord with Göllner’s [14] findings, and that, in the overall, the pattern of mandibular nerve arborization is as outlined by Göllner [14] (see his fig. 12, depicting an infant Gorilla sp.) and by Aziz et al. [20] (see their fig. 11, depicting the pattern in humans). Importantly, our dissections pointed out that the pattern of arborization of the mandibular nerve in the chimpanzee is nearly identical to its form in humans but the sizes and proportions of the mandibular nerve itself, its divisions and their branches are significantly different. The entire mandibular nerve complex of the chimpanzee is more robust (wider, thicker, bulkier, and longer nerves) than in humans. The same is true regarding the maxillary artery and its branches (Aziz et al., work in progress).

DISCUSSION
The mandibular division of the trigeminal nerve supplies a substantial group of head muscles which facilitate mastication, non-verbal communication, respiration, panting, vocal modulation (and speech, in humans), deglutition
(tensor veli palatini, mylohyoid and anterior digastric), sound conduction and its modulation (tensor tympani). These functional groups of muscles, that are subparts of the “trigeminal musculature”, are located around and supplied by the mandibular nerve tree. The muscular branches of this system arborise in several contiguous spaces of the head region: 1) the masseteric “fossa”; 2) the temporal fossa; 3) the infratemporal fossa; 4) the pterygoid fossa; and 5) the auditory tube. These spaces accommodate almost all the above-mentioned functional groups of muscles which are derived from the mesoderm of the mandibular arch. In previous studies of primates the “trigeminal musculature” has been exposed and described on a piecemeal basis resulting in a collage of disjointed narratives that are not always easy to connect [29, 30, 48]. It is thus instructive to summarize and critique the dissection protocols that have been employed so far to expose the territories which house parts of the “trigeminal musculature”. The dissection protocols described below refer to humans; however, we also indicate their use in exposing parts of the masticatory space in the nonhuman primates.
-
The Lateral Approach. This is the best-described approach; it is a prominent part of most contemporary human anatomy dissection guides [62, 65-67], atlases [61, 63, 68-72] and leading text books [73, 74]. This procedure of exposing the superficial parts of the infratemporal fossa is a refinement of the method that was originally used by Gray [74] whose book also includes instructions on the sequential process of dissection to expose the contents of the infratemporal fossa. Essentially this protocol involves the osteotomy of the zygomatic arch and much of the mandibular ramus [68]. It allows us to observe the outer surfaces of the intermediate parts of the buccal, the lingual, and the inferior alveolar nerves; the maxillary artery (and vein) and its deep temporal and buccal branches (tributaries).
In order to view the main mandibular stump of the nerve and branches of its “anterior division”, the dissection guides suggest the complete removal of the lateral pterygoid by blunt dissection. The removal of this pivotal lateral pterygoid, in addition to the removal of its own complex of nerves and vessels, dispenses with the very myological landmark without which cartography of this space is problematic. The lateral pterygoid heads are critical to the mapping of the mandibular nerve tree, the maxillary artery and its branches, and the organization of the maxillary vein and the pterygoid venous plexus. The removal of this muscle scrambles the morphology of all these critical structures. Unfortunately, this method has become the standard operating procedure in introductory anatomy courses offered to freshman medical and dental students [68]. The drawbacks of this approach are numerous. It precludes the direct observation of: the mylohyoid and the anterior digastric muscles (and their neurovascular structures); the deeper aspects of the medial pterygoid and the tensor veli palatini; the otic ganglion; nerves and vessels to the medial pterygoid, the tensor veli palatini, and the tensor tympani. However, deep dissection of the infratemporal fossa does expose the auriculotemporal nerve and the middle meningeal artery (which enters the foramen spinosum), as well as the sphenomandibular and the stylomandibular ligaments and their attachments.
This dissection protocol has also been used by surgeons to remove tumors of the deep infratemporal fossa and other contiguous spaces [75, 76]. It is also employed by dental anatomists to instruct students in the optimal application of inferior alveolar nerve block to anaesthetise the mandible and its teeth for dental procedures [69]. This method of exposure of the infratemporal fossa has been used in other primates, e.g., the chimpanzee [14, 15], the gorilla [14], the orangutan [77-79], the baboon [15, 21] and the rhesus monkey and its relatives [16, 21, 81, 82]. Skinner and Aziz [82] have written a critical analysis of this approach as applied to the Cercopithecoidea. - The Medial Approach. This protocol is designed to bring to full view the medial aspect (i.e the deep view) of the lateral pterygoid, the medial pterygoid, and the tensor veli palatini. This method is also used to expose the main trunk of the mandibular nerve and its divisions and their proximal branches. Also exposed are the meningeal branch, the nerves to the tensor veli palatini, the tensor tympani, and the otic ganglion. The bifurcation of the mandibular nerve into its two major divisions, the anterior and the posterior, can also be appreciated using this approach. One can also observe the origins of the proximal branches of each of these divisions. The auriculotemporal nerve and its associated middle meningeal artery can also be clearly seen. The medial approach necessarily involves hemisection of the head and the osteotomy of areas surrounding the foramina ovale and spinosum 21, 57, 72, 80, 83-91. Göllner 14 used this approach in the chimpanzee and the gorilla. Schwartz and Huelke 16 and Gaspard et al. 83, 84 applied it to the rhesus monkey and its relatives. This approach is appropriate to observe the deep aspects of the lateral pterygoid, the medial pterygoid, and the tensor veli palatini. It is also the best way to view the proximal parts of the mandibular nerve and its branches, and the otic ganglion. However, neither the outer aspects of all of these structures nor their connections with other parts of the “trigeminal musculature” can be appreciated, although one can improve prospects by combining the lateral and medial approaches [82].
- The Superior Approach. This was originally devised by Pinto [55] specifically to test the anatomical basis of the so-called Costen’s Syndrome, i.e. the projection of TMJ pain into the tympanic cavity. A prerequisite of this protocol is craniotomy including the removal of the brain and the meninges. Once the floor of the middle cranial fossa has been exposed, the greater wing of the sphenoid is removed by careful drilling and sawing; this method has been further refined by Sugisaki et al. [56]. It brings into view the mandibular nerve as it leaves the middle cranial fossa to enter the roof of the infratemporal fossa and the bifurcation of the mandibular nerve into its anterior and posterior divisions. One can observe the arborization of the anterior division and chart the paths of its branches – the masseteric and the posterior and middle deep temporal nerves, the proximal parts of the buccal nerve (and branches to the lateral pterygoid - direct as well as those that arise from the buccal nerve). Most of these nerves arborize over the upper surface of the superior lateral pterygoid head. The buccal nerve is directed anterolaterally before it enters the cleft between the two lateral pterygoid venters. This approach also exposes the dorsal aspects of the TMJ disc and the bilaminar tendon that is located posterior to the joint. Abe [104] combined this and the lateral approach to investigate the attachments of the lateral pterygoid head to the mandibular condyle and their relationships with the contiguous branches of the mandibular nerve. To our knowledge, this method has not been used to investigate parts of the “trigeminal musculature” or their nerve supplies in non-human primates.
- The Posterior (or inferior) Approach. This method involves the removal of the entire floor of the oral cavity, including the tongue and the posterior part of the pharynx. It generates a panoramic view of the pterygoid muscles and the muscles of the soft palate in the retro-choanal area [71, 72]. However, it is not possible to observe most of the mandibular nerve tree or the branches of the maxillary artery. To our knowledge, this method has also not been used in non-human primates.
- Lateral Pterygoidectomy. This en bloc approach has traditionally involved the removal of the mandibular condyle, TMJ disc, the lateral pterygoid heads, the mandibular nerve and its proximal parts/branches, and most of the maxillary vasculature. The major purposes of this procedure have been to assess the number of venters of the lateral pterygoid, determine the degree of their separation, and establish the true nature of the attachment of the TMJ disc with the superior lateral pterygoid head [60, 92-110]. Another objective of this method has been the need to establish the nerve supply to the lateral pterygoid heads [20, 57, 59, 111-113]. This protocol has recently been augmented to include the medial pterygoid and the tensor veli palatini, in addition to the lateral pterygoid heads [59]. Akita et al. [48] removed the lateral pterygoid and the temporalis along with the mandibular nerve (including its anterior division) to study the morphological and ontogenetic connections between these muscles (or their subparts) [48, 49]. Lashley [114] has used this method - described in detail by Aziz et al. [59] - to extract the lateral and medial pterygoid and the tensor veli palatini (along with the entire mandibular nerve tree) from the head of the chimpanzee. She has also comprehensively described the nerve supply to these muscles in the chimpanzee.
It can thus be said that the methods of exposing the “trigeminal musculature” described above provide fragmentary information regarding the components of the mandibular arch developmental field. The en bloc method of removing the entire mandible along with all (except the tensor tympani) derivatives of the mandibular arch pre-muscle plate which we have described in the present work opens new possibilities to examine these derivatives from an integrated, developmental perspective. Tomo and his associates have provided promising possibilities of arriving at a sound, ontogenetic classification of the muscles supplied by the mandibular nerve tree [17-19]. Specifically, they used detailed dissection of the branches of the mandibular nerve which supply the medial and lateral pterygoid muscles of the dog in order to show their separate nerve supplies and origins from the mandibular arch plate. They established that these muscles arise from distinct blocks of the ventromedial anlage [35, 36]. Shimokawa and Akita and their colleagues have used this approach to begin a fresher and more satisfactory ontogenetic classification of human masticatory muscles [29, 30, 48, 49]. However, these investigations have so far been limited to the masseter-temporalis-lateral pterygoid complex. Our approach permits a more comprehensive and holistic examination of the entire “trigeminal musculature” complex. This method will facilitate a definitive anatomical test of the hypothesis regarding ontogeny and phylogeny of the masticatory muscles as proposed by Edgeworth [35, 36] and, more recently, by Diogo et al. [10].
Schwartz and Huelke [16] (see their fig. 7), Tomo et al. [18] (see their fig. 8) and Aziz et al. [20] (see their fig. 11) have shown the branching of the mandibular nerve and the muscular targets of specific branches as viewed in coronal sections of the infratemporal fossae of the monkey, the dog, and the human, respectively. Such mapping helps to conceptualize the origins and migration of specific masticatory muscles from the subdivisions of the mandibular arch plate.
In deference to the attenuated contemporary medical curriculum [115] leading new anatomy text books tend to take an excessively reductionist approach to the description of the masticatory muscles. For example, the masseter and the medial pterygoid muscles are described as being three and two layered, respectively. Similarly, the temporalis and the lateral pterygoid are shown as singular or biventral muscles, respectively [64-73]. However, a large body of research from the last half century has conclusively shown that these muscles, and particularly the temporalis and masseter, are constituted of several extra-and intra-muscular subparts that play subtle roles in generating graded movements during the closing phase of the masticatory cycle. Detailed anatomical [21-22, 24-27, 29, 30, 83, 86, 116, 117] and physiological studies of the masseter muscle of humans and nonhuman primates [87, 118, 119] attest to its actual anatomical and functional heterogeneity. Similar evidence of anatomical [17, 21, 22, 25, 29, 30, 44, 45, 47, 120] and physiological complexity [119, 121] of the temporalis exists in published literature. The medial pterygoid [21, 22, 84, 85, 121-124] and the lateral pterygoid [20, 48, 58, 59, 84, 85, 95, 104, 121, 125-127] also exhibit formal as well as functional diversity.
Hannam and McMillan [128] have made the following statement regarding our knowledge of the masticatory muscles: “most of these muscles are not simple; they are multipennate, complexly layered, and divided by aponeuroses; this arrangement supplies substantial means for differential contraction.” Therefore, precise mapping of the trajectories of the branches of the mandibular nerve which supply each one of the masticatory muscles becomes a significant issue because “there is circumstantial evidence for intramuscular partitioning based in part on innervation by primary muscle nerve branches” [128]. This information is critical for obtaining reliable results in EMG studies of the masticatory muscles.
Recently, there has been considerable controversy regarding the following aspects of the masticatory muscles: 1) the distinct subparts of each named muscle; 2) the existence of muscle slips/bundles between the masseter and the temporalis (e.g. the “maxillomandibularis”, the “zygomaticomandibularis”); 3) the existence of muscle slips (e.g., “discal muscle slips”) of the masseter and the temporalis that are attached to the TMJ disc; 4) claims of the existence of a so-called “sphenomandibularis” muscle located deep to the temporalis; and 5) the status of the “pterygoid proprius” and the “pterygo-spinous” muscles which have been found deep to the lateral pterygoid. These issues may now be more satisfactorily resolved using the integrated approach such as the one made possible by our protocol [25, 29, 30, 45-49, 129]. For example, Akita et al. [48] used a method to show that the lateral pterygoid and various muscle slips between it, the temporalis and the TMJ capsule are commonly derived from the “midmedial part of the anlage of the masticatory muscles” [36, 35].
The precise cartography of the masticatory muscles and their individual nerve trees is critical to our comprehension of their functional and clinical applications. For instance, it is now well-established that each compartment of each masticatory muscle is involved in performing a particular function - “functional heterogeneity”, see Herring et al. [87] - during the masticatory cycle [21, 58, 124, 130-132]. Aziz et al. [20] have critiqued the claims, based on electromyography, of antagonistic behavior of the lateral pterygoid heads during the masticatory cycle. That is, the lateral pterygoid heads are probably antagonistic in some of their actions, at least. But EMG data presented in the literature so far are open to debate because the physiological studies were not based on sufficient prior anatomical analyses. Credible claims can only be made when electromyographic studies dovetail meticulously detailed mapping of the investigated masticatory muscle itself (including its subparts) and its nerve tree [18, 20, 58].
Ultimately, the question as to whether the two lateral pterygoid heads of humans and/or other mammals are controlled by isolated pools of motoneurons in the trigeminal nucleus can only be definitively resolved by sophisticated neural topography or mapping such as used by Graf et al. [133]. The pterygoid heads also play a significant role (in addition to other related anatomical structures) in movements that facilitate speech in humans. If it can be shown that the two heads have evolved to perform nuanced synchronous and asynchronous actions during the masticatory cycle, we may discover a morphological link in the origin of speech in derived hominoids.
Numerous investigations on mammalian models (e.g. rat, guinea pig, cat, and the rhesus monkey) have attempted to test Edgworth’s [35, 36] and Lightoller’s [134] ontogenetic and phylogenetic classification schemes of the mandibular and hyoid muscles of ancestral and derived chordates (see Diogo et al. [10] for a recent, updated overview of these phylogenetic and classification schemes). Amongst these tests is the use of sophisticated contemporary neural topographical methods involving the retrograde transport of various tracers. Mapping of the motor trigeminal nucleus shows that distinct motoneuron pools of the masseter and temporalis are located dorsolaterally while those of the pterygoids (“jaw closing” muscles) are placed ventromedially [135, 136]. The motoneurons of the anterior digastric and the mylohyoid are found in the ventromedial part of the nucleus. Rokx and van Willingen [136] write that the controls of the lateral pterygoid - which, they observe “contracts in concert with closing muscles in the bite proper and the power stroke but is also active during jaw opening” - are located in an “intermediate (position) between opening and closing motoneurons”.
In summary, comparative morphological, ontogenetic and phylogenetic, observations on extant mammalian genera show a remarkable concordance. Much of the evidence points to the organization of motoneurons of numerous head muscles to reflect “location according to common function” within their respective nuclei. Yet, Akita et al. [48] have found that we need a much more meticulous mapping of the mandibular nerve tree within individual masticatory muscles in order to fill significant gaps in our knowledge of the trigeminal musculature which includes many muscles which are individually partitioned such that each muscle acts in the manner of several distinct muscles (see above).
The method to extricate the entire “trigeminal musculature” as outlined in this report also promises an additional dividend: the measurement of the phenotypic distance between the “trigeminal musculature” of the chimpanzee and humans. We now possess substantially complete genomic maps of both genera [137, 138]. For instance, Sibley et al. [139] have estimated that humans and the chimpanzee share 98.6% of their genetic material. However, despite this close genetic relationship, the head structures of these genera show obvious phenotypic differences, e.g., the chimpanzee skull is considerably more robust, more prognathous and features exceptionally deep temporal, masseteric and infratemporal fossae, a more robust dentition, including prominent canines, and a massive scale of all masticatory (and related) muscles; this increased muscle mass is reflected in the larger dimensions of the mandibular nerve and its branches (see above). Stedman et al. [140] proposed that a “gene encoding the predominant myosin heavy chain (MYH) expressed in these (i.e. the pongids) muscles was inactivated by a frame shifting mutation after the lineages leading to human and chimpanzee diverged.”
Given that the chimpanzee is one of the most highly protected primates, its tissues are now not so readily available for anatomical research. Therefore, the development of a method that permits the most comprehensive study of the “trigeminal musculature” may be the most economical use of any chimpanzee cadaveric tissues that do become available for anatomical studies of this complex and functionally very important musculature.
ACKNOWLEDGEMENTS
This study was supported by funds from Howard University College of Dentistry, and the Department of Anatomy of the Howard University College of Medicine (James H. Baker, Ph.D., Chairman). The chimpanzee specimen (HUC 102) was acquired from the Yerkes Regional Primate Research Center (Emory University, Atlanta, Ga.), and we are very grateful to the Associate Director for Research Resources, Dr. James Else and to Ms. Kay Lee Summerville, Senior Program Associate [Yerkes Base Grant RR00165] for their kind assistance. We also thank Mr. Greg Robertson (Photography), Ms. Lynette Thompson (Desktop Publishing), Mr. James Ferguson (Lab Assistance) and especially to Mr. Stafford Battle (Computer Graphics) for their contributions.