- Home
- About Journals
-
Information for Authors/ReviewersEditorial Policies
Publication Fee
Publication Cycle - Process Flowchart
Online Manuscript Submission and Tracking System
Publishing Ethics and Rectitude
Authorship
Author Benefits
Reviewer Guidelines
Guest Editor Guidelines
Peer Review Workflow
Quick Track Option
Copyediting Services
Bentham Open Membership
Bentham Open Advisory Board
Archiving Policies
Fabricating and Stating False Information
Post Publication Discussions and Corrections
Editorial Management
Advertise With Us
Funding Agencies
Rate List
Kudos
General FAQs
Special Fee Waivers and Discounts
- Contact
- Help
- About Us
- Search




The Open Ornithology Journal
(Discontinued)
ISSN: 1874-4532 ― Volume 13, 2020
Molecular Systematics, Taxonomy and Distribution of the Pyrrhura Picta–Leucotis Complex
Thomas Arndt1, *, Michael Wink2
Abstract
Introduction:
The relationships within the Pyrrhura species complex are partly unresolved. In this study, a comprehensive phylogenetic analysis of the Pyrrhura picta-leucotis complex was carried out, covering all species except P. subandina.
Material and Methods:
We made a morphological analysis of 745 preserved specimens of all the taxa in different museums. Nucleotide sequences of the mitochondrial cytochrome gene were generated and used to reconstruct a molecular phylogeny of Pyrrhura.
Results and Discussion:
Our results show that the complex is divided into 6 main groups comprising 15 species. P. dilutissima, regarded up to now as a subspecies of P. peruviana, acquires species status and three new subspecies are described. We also provide evidence that P. roseifrons is a paraphyletic group, indicating the existence of probably 3 lineages of which 2 deserve species status.
Article Information

Identifiers and Pagination:
Year: 2017Volume: 10
First Page: 53
Last Page: 91
Publisher Id: TOOENIJ-10-53
DOI: 10.2174/1874453201710010053
Article History:
Received Date: 09/09/2016Revision Received Date: 05/01/2017
Acceptance Date: 15/02/2017
Electronic publication date: 19/05/2017
Collection year: 2017
open-access license: This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International Public License (CC-BY 4.0), a copy of which is available at: (https://creativecommons.org/licenses/by/4.0/legalcode). This license permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
* Address correspondence to this author at the Brueckenfeldstr. 28, 75015 Bretten, Germany; Tel: (+49) 7252 97073-10; Fax: (+49)-7252-97073-25; E-mail: thoarndt@aol.com
| Open Peer Review Details | |||
|---|---|---|---|
| Manuscript submitted on 09-09-2016 |
Original Manuscript | Molecular Systematics, Taxonomy and Distribution of the Pyrrhura Picta–Leucotis Complex | |
1. INTRODUCTION
The genus Pyrrhura, especially the species-level systematics of the Pyrrhura picta-leucotis complex, has long been problematic. While Forshaw and Cooper [1Forshaw JM, Cooper WT. Parrots of the world. 3rd (rev) ed. London: Blandford Press 1989.], Collar [2Collar NJ. Psittacidae (Parrots). In: del Hoyo J, Elliott A, Sargatal J, Eds. Handbook of the birds of the world. Lynx Edicions, Barcelona 1997; pp. 280-477.] and Juniper and Parr [3Juniper T, Parr M. Parrots: A guide to the parrots of the world. Robertsbridge, UK: Pica Press 1998.] list two species, Joseph [4Joseph L. Beginning an end to 63 years of uncertainty: The Neotropical parakeets known as Pyrrhura picta and P. leucotis comprise more than two species. Proc Acad Nat Sci Philadelphia 2000; 150: 279-92., 5Joseph L. Geographical variation, taxonomy and distribution of some Amazonian Pyrrhura parakeets. Ornith Neotrop 2002; 13: 337-63.] was the first to split the complex, and additionally he described two new species, Pyrrhura snethlageae and P. peruviana. Ribas, et al. [6Ribas CC, Joseph L, Miyaki CR. Molecular systematics and patterns of diversification in Pyrrhura (Psittacidae) with special reference to the picta-leucotis complex. Auk 2006; 123: 660-80.
[http://dx.doi.org/10.1642/0004-8038(2006)123[660:MSAPOD]2.0.CO;2] ] provided the first genetic results and Arndt [7Arndt T. Anmerkungen zu einigen Pyrrhura-Formen mit der Beschreibung einer neuen Art und zweier neuer Unterarten. Papageien 2008; 21: 278-86.] added a further new species, P. parvifrons. While the Zoological Nomenclature Resource (www.zoonomen.net) mainly follows Ribas, et al. [6Ribas CC, Joseph L, Miyaki CR. Molecular systematics and patterns of diversification in Pyrrhura (Psittacidae) with special reference to the picta-leucotis complex. Auk 2006; 123: 660-80.
[http://dx.doi.org/10.1642/0004-8038(2006)123[660:MSAPOD]2.0.CO;2] ] in the species identification, Del Hoyo and Collar [8Del Hoyo J, Collar NJ. HBW and BirdLife International Illustrated checklist of the birds of the World. Lynx Edicions, Barcelona: Non-Passerines 2014; Vol. 1.] in their new checklist of the birds of the world accept Arndt’s revision in full, which is based on the examination of museum specimens. In a recent paper, Gaban-Lima and Raposo [9Gaban-Lima R, Raposo MA. The status of three little known names proposed by Miranda-Ribeiro (1926) and the synonymization of Pyrrhura / snethlageae Joseph & Bates, 2002 (Psittaciformes: Psittacidae: Arinae). Zootaxa 2016; 4200: 192-200.
[http://dx.doi.org/10.11646/zootaxa.4200.1.10] [PMID: 27988647] ] showed that Pyrrhura pallescens is a valid senior name for P. snethlageae.
Joseph [5Joseph L. Geographical variation, taxonomy and distribution of some Amazonian Pyrrhura parakeets. Ornith Neotrop 2002; 13: 337-63.] provided the first excellent characterization of the taxa of the picta-leucotis group in the Amazon basin. However, Arndt [7Arndt T. Anmerkungen zu einigen Pyrrhura-Formen mit der Beschreibung einer neuen Art und zweier neuer Unterarten. Papageien 2008; 21: 278-86.] pointed out that the criteria used by Joseph [5Joseph L. Geographical variation, taxonomy and distribution of some Amazonian Pyrrhura parakeets. Ornith Neotrop 2002; 13: 337-63.] to characterize various forms and the colouring of the breast feathers were insufficient. The range of variation within the taxa is considerable. Coloration of the individual breast feathers and the breast in total is variable among individuals of the forms investigated, and therefore can only be used in combination with other characters to determine the various taxa. Nonetheless the taxa as circumscribed by Joseph are corroborated by our examination of the specimen material. Ribas, et al. [6Ribas CC, Joseph L, Miyaki CR. Molecular systematics and patterns of diversification in Pyrrhura (Psittacidae) with special reference to the picta-leucotis complex. Auk 2006; 123: 660-80.
[http://dx.doi.org/10.1642/0004-8038(2006)123[660:MSAPOD]2.0.CO;2] ] were able to support the validity of some species by molecular genetic means. However, this work demonstrated the limits of DNA analysis for the picta-leucotis group. Thus, for example, no clear genetic demarcation could be found between P. snethlageae and amazonum-microtera or between parvifrons and peruviana, although these taxa very clearly differ in morphology. This situation is not unusual and occurs in other young species complexes [10Rheindt FE, Székely T, Edwards SV, et al. Conflict between genetic and phenotypic differentiation: the evolutionary history of a lost and rediscovered shorebird. PLoS One 2011; 6(11): e26995.
[http://dx.doi.org/10.1371/journal.pone.0026995] [PMID: 22096515] , 11Filardi CE, Smith CE. Molecular phylogenetics of monarch flycatchers (genus Monarcha) with emphasis on Solomon Island endemics. Mol Phylogenet Evol 2005; 37(3): 776-88.
[http://dx.doi.org/10.1016/j.ympev.2005.02.007] [PMID: 16291094] ]. The most probable explanation for the picta-leucotis complex is that the splitting up of populations and taxa is very recent and reproductive isolation is still incomplete. In such situations, it is possible that morphological differences are more apparent than genetic ones (M. Schaefer, in litt.). Our molecular phylogeny research, which includes many taxa than previous studies, highlights this problem and shows that only a combination of phylogenetic, morphological, ecological and ethological data allow more precise taxonomic judgments.
Joseph [5Joseph L. Geographical variation, taxonomy and distribution of some Amazonian Pyrrhura parakeets. Ornith Neotrop 2002; 13: 337-63.] already pointed out that the use of the biological species concept [12Mayr E. Systematics and the origin of species. Cambridge, Mass: Harvard Univ. Press 1942.] is problematic in the picta-leucotis complex. This applies likewise for the morphological or phylogenetic species concepts. We also have noted Gill’s “new null hypothesis for species designation” [13Gill FB. Species taxonomy of birds: Which null hypothesis? Auk 2014; 131: 150-61.
[http://dx.doi.org/10.1642/AUK-13-206.1] ], which, in the absence of any natural tests of reproductive isolation, considers genetically and phenotypically distinct taxa as full species, but agree with Joseph [5Joseph L. Geographical variation, taxonomy and distribution of some Amazonian Pyrrhura parakeets. Ornith Neotrop 2002; 13: 337-63.], that probably the complex is best characterized if it is regarded from the viewpoint of the Comprehensive Biologic Species Concept for birds [14Johnson NK, Remsen JV Jr, Cicer C. Resolution of the debate over species concepts in ornithology: A new comprehensive biologic species concept. Proc Int Ornithol Congr. 1470-82.]. This considers avian species as systems of populations representing essentially monophyletic, genetically cohesive and genealogically concordant lineages of individuals on independent evolutionary trajectories [5Joseph L. Geographical variation, taxonomy and distribution of some Amazonian Pyrrhura parakeets. Ornith Neotrop 2002; 13: 337-63.].
The method used by us for assigning species rank by Tobias, et al. [15Tobias JA, Seddon N, Spottiswoode CN, Pilgrim JD, Fishpool LD, Collar NJ. Quantitative criteria for species delimitation. Ibis 2010; 152: 724-46.
[http://dx.doi.org/10.1111/j.1474-919X.2010.01051.x] ] and Helbig, et al. [16Helbig AJ, Knox AG, Parkin DT, Sangster G, Collinson M. Guidelines for assigning species rank. Ibis 2002; 144: 518-25.
[http://dx.doi.org/10.1046/j.1474-919X.2002.00091.x] ] is based on the BSC. This is especially problematic for allopatric taxa, as without geographical contact reproductive isolation cannot be tested directly, which can lead to subjective results [12Mayr E. Systematics and the origin of species. Cambridge, Mass: Harvard Univ. Press 1942., 17Brown WL, Wilson EO. Character displacement. Syst Zool 1956; 5: 49-64.
[http://dx.doi.org/10.2307/2411924] -19Zink RM, McKitrick MC. The debate over species concepts and its implications for ornithology. Auk 1995; 112: 701-19.]. However, Tobias, et al. [15Tobias JA, Seddon N, Spottiswoode CN, Pilgrim JD, Fishpool LD, Collar NJ. Quantitative criteria for species delimitation. Ibis 2010; 152: 724-46.
[http://dx.doi.org/10.1111/j.1474-919X.2010.01051.x] ] point out that this does not need to be fatal as arbitrariness can be minimised using direct comparisons with related sympatric species [14Johnson NK, Remsen JV Jr, Cicer C. Resolution of the debate over species concepts in ornithology: A new comprehensive biologic species concept. Proc Int Ornithol Congr. 1470-82., 20Winker K. Subspecies represent geographically partitioned variation, a goldmine of evolutionary biology, and a challenge for conservation. Ornithol Monogr 2010; 67: 6-23.
[http://dx.doi.org/10.1525/om.2010.67.1.6] ].
We present in this work the most detailed distributional information to date, particularly for the taxa represented in the Amazon basin and Peru, which in part differ considerably from the known range descriptions (Appendix 1, Map 1 -5
-5 ). This was possible through the combination of museum specimen material and sightings in the wild, for the most part published on the Internet. However, the failure rate in the identification of species/subspecies appearing for Internet data is relatively high. Therefore, only those records, which were clearly supported by photos or detailed descriptions are considered in the maps.
). This was possible through the combination of museum specimen material and sightings in the wild, for the most part published on the Internet. However, the failure rate in the identification of species/subspecies appearing for Internet data is relatively high. Therefore, only those records, which were clearly supported by photos or detailed descriptions are considered in the maps.
The maps of the various taxa in the Amazon basin reveal their largely parapatric distributions and their relatively strict confinement to the diverse river systems Fig. (1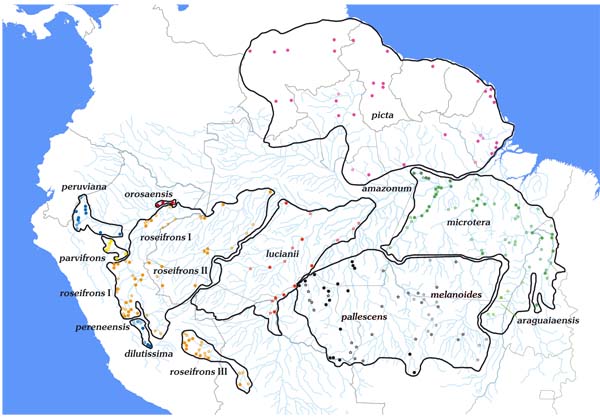 ). Thus one finds amazonum in the lower catchment area of the Rio Tapajós, Rio Xingu and Rio Tocantins/Rio Araguaia, lucianii along the middle Rio Amazonas, Rio Madeira and Rio Purus and roseifrons in the area of the upper Amazon/Río Ucayali, Rio Jurua and Río Madre de Dios. Pallescens holds a special position by occupying the upper catchment area of the Rio Madeira, Rio Tapajós and Rio Xingu. This distribution is probably explained by the forest refugium theory, which posits that during the Pleistocene period isolated areas of forest produced new species. The work of Bush and Oliveira [21Bush MB, Oliveira PE. The rise and fall of the refugial hypothesis of amazonian speciation: A paleoecological perspective. Biota Neotrop 2006; 6(1)
). Thus one finds amazonum in the lower catchment area of the Rio Tapajós, Rio Xingu and Rio Tocantins/Rio Araguaia, lucianii along the middle Rio Amazonas, Rio Madeira and Rio Purus and roseifrons in the area of the upper Amazon/Río Ucayali, Rio Jurua and Río Madre de Dios. Pallescens holds a special position by occupying the upper catchment area of the Rio Madeira, Rio Tapajós and Rio Xingu. This distribution is probably explained by the forest refugium theory, which posits that during the Pleistocene period isolated areas of forest produced new species. The work of Bush and Oliveira [21Bush MB, Oliveira PE. The rise and fall of the refugial hypothesis of amazonian speciation: A paleoecological perspective. Biota Neotrop 2006; 6(1)
[http://dx.doi.org/10.1590/S1676-06032006000100002] ] and Haffer and Prance [22Haffer J, Prance GT. Climatic forcing of evolution in Amazonia during the Cenozoic: On the refuge theory of biotic differentiation. Amazoniana 2001; 16: 579-608.] as well as Prance [23Prance GT. A Review of the phytogeographic evidences for Pleistocene climate changes in the Neotropics. Ann Mo Bot Gard 1982; 69: 594-624.
[http://dx.doi.org/10.2307/2399085] ] and Haffer [24Haffer J. Avian speciation in tropical South America. Cambridge: Nuttall Ornithological Club, Harvard University 1974.] discuss these forest refugia, showing that in parts of current distribution of P. pallescens such refugia are likely to be found.
The area over which the picta-leucotis occurs is huge, and between its many constituent taxa there are large gaps which may or may not be real. Nonetheless, there are a few localities where the ranges of different taxa have been shown to be parapatric. Thus pallescens and amazonum meet in the outermost north-east Mato Grosso in the area between Confresa and Alo Brasil for a few kilometres, picta and amazonum on the Río Trombetas, and roseifrons comes into contact in Peru with lucianii on the Río Orosa and Madre Selva as well as with parvifrons on the Río Cushabatay. The same is true for pallescens and lucianii in the area of Porto Velho and roseifrons and dilutissima in the Peruvian Río Perene valley.
Relatively small isolated populations have developed in the Peruvian Andes in the larger river valleys. These are highly restricted in their extent of occurrence, have a movement radius of only a few kilometres and confined to certain altitudes. Klauke, et al. (in press) [25Klauke N, Schaefer HM, Bauer M, Segelbacher G. The influence of natural habitat structure and habitat disturbance on the fine-scale genetic structure of an endemic Neotropical parrot. Mol Ecol submitted] have demonstrated this for Pyrrhura orcesi and Arndt and Gonzales Pinedo [26Arndt T, Gonzales Pinedo H. Die Rotschwanzsittiche in der Cordillera Escalera, Peru. Papageien 2013; 26: 204-7.] for P. parvifrons. Klauke, et al. [25Klauke N, Schaefer HM, Bauer M, Segelbacher G. The influence of natural habitat structure and habitat disturbance on the fine-scale genetic structure of an endemic Neotropical parrot. Mol Ecol submitted] point out that this strong site fidelity greatly affects population development and the conservation status of threatened (sub) populations, as the birds are unable to move to other areas, even if these seem easily accessible and are just a few hundred metres higher or lower. Field observations by TA, who visited practically all the larger Peruvian and Brazilian populations between 2008 and 2015, suggest that this behaviour is characteristic for all Peruvian representatives of Pyrrhura in the Andes region.

This unusually low dispersal range can be explained by certain typical behavioural patterns. According to previous observations (e.g. [27Girão W, Campos A, Albano C. Das Schutzprojekt für den Salvadori-Weißohrsittich. Papageien 2008; 21: 29-32.], field observations by TA) most if not all Pyrrhura representatives have traditional roosting/nesting trees, which have been used for decades by small groups of up to 10 birds and probably make them very conservative in their dispersive abilities. The birds mainly use living trees such as Albizia polycephala and Inga sp [27Girão W, Campos A, Albano C. Das Schutzprojekt für den Salvadori-Weißohrsittich. Papageien 2008; 21: 29-32.], and the absence of these trees appears to be a significant limiting factor in the ability of birds to occupy otherwise seemingly suitable habitat.
The specific habitat requirements of most of the Peruvian Pyrrhura species are still incompletely known, but it seems that many populations require intact mountain rain forest with tall tree species. They forage at lower elevations in more fragmented areas at certain times of the year, but are generally absent from forested areas, which have been affected by human activities. This suggests special habitat requirements, which result in localised occurrence.
Moreover, Klauke et al. (in press) [25Klauke N, Schaefer HM, Bauer M, Segelbacher G. The influence of natural habitat structure and habitat disturbance on the fine-scale genetic structure of an endemic Neotropical parrot. Mol Ecol submitted] have shown in the case of Pyrrhura orcesi that even small geographical barriers such as valleys, which could be easily overflown, create obstacles that limit the dispersal of the birds. They have demonstrated that geographical barriers promote genetic divergence in quite small spaces.
The populations north and south of the Amazon basin occur allopatrically and are found, with the exception of P. leucotis, in isolated mountain areas with differing habitats and ecological as well as biogeographical histories, which have favoured their genetic divergence. This can be seen in the molecular phylogeny data for eisenmanni, caeruleiceps, emma, pfrimeri and griseipectus.
Here, for the first time we examine the Pyrrhura picta-leucotis complex through the combination of genetic (sequences of mitochondrial cytochrome b) and morphological analyses. Adding biogeographic and ethological data enabled us to understand the species-level systematics of the different taxa more precisely as well as to propose taxonomical changes. In addition, new findings were gained on the distribution of the taxa.
2. MATERIALS AND METHODS
2.1. Phenotypic Analysis
Museum Specimens and Biometric Data
Altogether we were able to examine 745 specimens of all known species and subspecies of the Pyrrhura picta leucotis complex, of which 7 were available as photographs and 578 were measured (wing, tail and bill, 4 from living birds) from specimens curated in the following museums: American Museum of Natural History in New York (AMNH), Carnegie Museum of Natural History in Pittsburgh (CM), Colección Ornitológica Phelps in Caracas (COP), Field Museum of Natural History in Chicago (FMNH), Louisiana State University Museum of Natural Science in Baton Rouge (LSUMZ), Museo de Historia Natural “Javier Prado” de la UNMSM in Lima (MJPL), The Natural History Museum in Tring (NHM), Museu Goeldi in Belém (MPEG), Staatliches Museum für Tierkunde in Dresden (MTD), Museu de Zoologia da Universidade de São Paulo in São Paulo (MZUSP), Netherlands Centrum voor Biodiversiteit in Leiden (RMNH), Forschungsinstitut and Museum Senckenberg in Frankfurt am Main (SMF), Museu Nacional in Rio de Janeiro (UFRJ), United States National Museum in Washington (USNM), Universidade de São Paulo in São Paulo (USP), Museum für Naturkunde der Humboldt-Universität in Berlin (ZMB), and Zoologische Staatssammlung München (ZSM).
The colour terminology for describing the type specimens is based on Smithe [28Smithe FB. Naturalist’s color guide. New York: American Museum of Natural History 1975.].
Vocal Analysis
Differences in voice are often indicative of geographic and genetic isolation; vocalisations are not only used for determining the species, but above all to establish whether we are dealing with closely related allopatric speciation or not [29Alström P, Ranft R. The use of sounds in avian systematics and the importance of bird sound archives. Bull Bull Brit Orn Cl 2003; 123: 114-35.].
We used for this purpose a very simplified method to compare calls and highlight probable differences between those of different taxa, without claiming to prove them definitively.
Call sequences were taken either from xeno-canto.org and ibc.lynxeds.com or from our own collection, using Raven Pro 1.5 as the analysis program. Calls were selected on the basis of their comparability (homology); usually they were flock calls. A spectrogram was created in Raven Pro from these sound samples and was examined if it exclusively contained Pyrrhura calls. Other calls or noises were removed. Finally, the peak frequency was determined.
2.2. Taxon Sampling for DNA Studies
Blood and Tissue Samples
For this research, we included nucleotide sequences of 49 samples from GenBank. The sequence dataset was extended by 33 new sequences generated in our own laboratory (Appendix 2). The comprehensive dataset covers the entire picta-leucotis complex with the exception of P. subandina.
2.3. DNA Extraction, Amplification, Sequencing and Analysis
DNA was obtained from blood samples stored in a modified EDTA buffer at -20°C, in 80% ethanol, or dried on filter paper until processing in the Heidelberg laboratory. Total DNA was isolated using standard proteinase K (Merck, Darmstadt) and phenol/chloroform procedures [30Wink M, El-Sayed A-A, Sauer-Gürth H, Gonzalez J. Molecular phylogeny of owls (Strigiformes) inferred from DNA sequences of the mitochondrial cytochrome b and the nuclear RAG-1 gene. Ardea 2009; 97: 209-19.
[http://dx.doi.org/10.5253/078.097.0425] , 31Wink M, Sauer-Gürth H. Phylogenetic relationships in diurnal raptors based on nucleotide sequences of mitochondrial and nuclear marker genes. In: Chancelor RD, Meyburg B-U, Eds. Raptors Worldwide. Berlin: WWGBP 2004; p. 483498.].
We amplified the mitochondrial cytochrome b gene (> 900 nucleotides; nt) as an informative marker gene which has been used by us before for a phylogenetic reconstruction of many other bird taxa. The PCR amplifications were performed in 50 µl reaction volumes containing 1 × PCR buffer (Bioron, Ludwigshafen), 100 µM dNTPs, 0.2 units of Taq DNA polymerase (Bioron, Ludwigshafen), 200 ng of DNA and 5 pmol of primers.
Thermal cycling was carried out under the following conditions: 5 min at 94°C, followed by 35 cycles of 40 s at 94°C, 40 s at 52.0°C, 1 min at 72°C and a final extension at 72°C for 10 min. PCR products were precipitated with 4 M NH4Ac and ethanol (1:1:6) and a centrifugation for 15 min (13,000 rpm).
Sequencing was performed using the ABI 3730 automated capillary sequencer (Applied Biosystems, CA, USA) with the ABI Prism Big Dye Terminator Cycle Sequencing Ready Reaction Kit 3.1 (carried out by STARSEQ GmbH, Mainz, Germany). For sequencing, the same primers were used as for the initial PCR amplifications.
2.4. Phylogenetic Analyses
The nucleotide sequences were aligned manually with BioEdit version 7.0.9.0. No internal stop codons or frame-shifts were observed in the sequences, which were translated entirely by using the chicken mitochondrial code.
Phylogenetic trees were reconstructed using the Maximum Likelihood (ML) algorithm in MEGA version 7 [32Kumar S, Stecher G, Tamura K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol Biol Evol 2015. submitted
[PMID: 27004904] ] with related parrot species as outgroups. The percentage of trees in which the associated taxa clustered together is shown next to the branches. Initial tree(s) for the heuristic search were obtained automatically by applying Neighbor-Join and Bio NJ algorithms to a matrix of pairwise distances estimated using the Maximum Composite Likelihood (MCL) approach, and then selecting the topology with superior log likelihood value. A discrete Gamma distribution was used to model evolutionary rate differences among sites (5 discrete Gamma categories {+G, parameter = 0.4968}). The rate variation model allowed for some sites to be evolutionarily invariable ({+I}, 33.6773% sites). The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. The analysis involved 84 nucleotide sequences. Codon positions included were 1st+2nd+3rd+Noncoding. There were a total of 1140 positions in the final dataset. Evolutionary analyses were conducted in MEGA7.
Sequence data have been submitted to GenBank (accession numbers listed in Appendix 2, taxon samplings).
2.5. Method for Assigning Species Rank
Practically all the taxa of our dataset show a parapatric or allopatric distribution. Because the phylogeny of the picta-leucotis complex reveals well-separated lineages, the “guidelines for assigning species rank” by Helbig et al. [16Helbig AJ, Knox AG, Parkin DT, Sangster G, Collinson M. Guidelines for assigning species rank. Ibis 2002; 144: 518-25.
[http://dx.doi.org/10.1046/j.1474-919X.2002.00091.x] ] as well as the ”quantitative criteria for species delimitation” established by Tobias et al. [15Tobias JA, Seddon N, Spottiswoode CN, Pilgrim JD, Fishpool LD, Collar NJ. Quantitative criteria for species delimitation. Ibis 2010; 152: 724-46.
[http://dx.doi.org/10.1111/j.1474-919X.2010.01051.x] ] provide a basis for a quantative delimitation of the species and subspecies in this complex. A parapatric taxon can be diagnosed using the Helbig system (e.g. showing at least two unique characters) but to meet this criterion it should not hybridise. Allopatric taxa must be clearly diagnosable by at least one character and the level of divergence must be equivalent to that between the two most closely related sympatric species.
Tobias et al. [15Tobias JA, Seddon N, Spottiswoode CN, Pilgrim JD, Fishpool LD, Collar NJ. Quantitative criteria for species delimitation. Ibis 2010; 152: 724-46.
[http://dx.doi.org/10.1111/j.1474-919X.2010.01051.x] ] use a quantitive system in which points are given as follows: for each minor character (a weak difference, e.g. a change in shade) 1, medium character (a clear difference reflected, e.g., by a distinct hue rather than different colour) 2, major character (a pronounced and striking difference in the colour or pattern of a body part, or in a measurement or vocalisation) 3, and exceptional difference (a radically different coloration or pattern) 4 points. In the evaluation system only three plumage characters, two biometric characters, two vocal characters and one behavioural or ecological character may be scored. The minimum number of points for species status is 7.
We have modified this point system and used it for all taxa, especially for those that are not clearly genetically defined in their group (see 3.10 taxonomic results), but take into account the demands of Helbig et al. [16Helbig AJ, Knox AG, Parkin DT, Sangster G, Collinson M. Guidelines for assigning species rank. Ibis 2002; 144: 518-25.
[http://dx.doi.org/10.1046/j.1474-919X.2002.00091.x] ] that in all allopatric taxa the level of divergence must be equivalent to that found in related sympatric species. In cases where a recognisable genetic difference exists between two taxa, we have weighted this with 1 to 4 points as one of the two possible biometric characteristics. For this evaluation, either bootstrap values or genetic distances can be used. Both result in about the same values. For bootstrap values the scoring would be: <80 = 0 point; 80-84 = 1 point; 85-90 = 2 points; 91-97 = 3 points; >97 = 4 points. We used genetic distances (uncorrected p distance) (Appendix 3), which describe the genetic differences more clearly. To determine the minimum and maximum distance values (1 = 100%) for the scoring we used the values of amazonum/pallescens (0.0052) and closely related Pyrrhura species pairs (molinae/frontalis: 0.0182; lepida/perlata: 0.0147; rhodocephala/hoffmanni: 0.0182). We scored with: < 0.0060 = 0 point; 0.0060-0.0089 = 1 point; 0.0090-0.0119 = 2 points; 0.0120-0.0149 = 3 points; > 0.0150 = 4 points. This system allows a considerable influence for genetic data, so that taxa with the maximum possible four points achieve already more than the half of the necessary score for species status.
In establishing biometric characters, we have limited ourselves to bill, wing and tail measurements and used the “Cohen’s d” value required by Tobias et al. [15Tobias JA, Seddon N, Spottiswoode CN, Pilgrim JD, Fishpool LD, Collar NJ. Quantitative criteria for species delimitation. Ibis 2010; 152: 724-46.
[http://dx.doi.org/10.1111/j.1474-919X.2010.01051.x] ].
As we are using a very simplified form of the vocal analysis, we have only allowed one point where differences in the peak frequency were established, so as not to over-value this character.
The assessment of ecological data as a species character is only partially helpful, as indicated also by Tobias et al. [15Tobias JA, Seddon N, Spottiswoode CN, Pilgrim JD, Fishpool LD, Collar NJ. Quantitative criteria for species delimitation. Ibis 2010; 152: 724-46.
[http://dx.doi.org/10.1111/j.1474-919X.2010.01051.x] ], because at least widespread Pyrrhura taxa such as amazonum, pallescens and roseifrons vary geographically in their microhabitat requirements. We have drawn not only on published information but also on the findings of our own fieldwork. In evaluating the species rank system we were very cautious and only awarded one point where the habitat use clearly indicated specialisation.
2.6. Cluster Analysis
To control and confirm the results of the species rank method we carried out a cluster analysis using 31 character groups (24 plumage and soft parts, 4 biometrics, 2 ecological, 1 allopatric/parapatric disjunction) in 91 characters. The results of the analysis are shown in (Fig. 2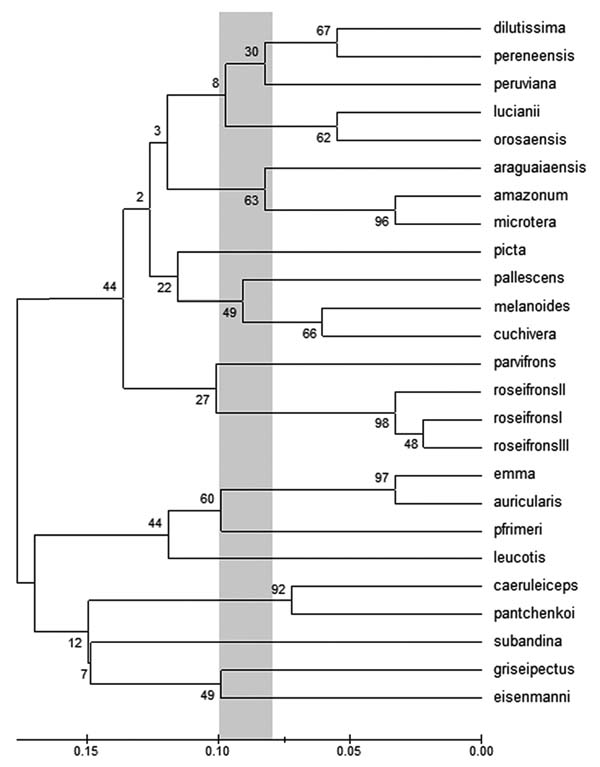 ).
).
 |
Fig. (2) Cluster analysis using 31 character groups (24 plumage and soft parts, 4 biometrics, 2 ecological, 1 allopatric/parapatric disjunction) and 91 characters. |
3. RESULTS AND DISCUSSION
3.1. Sequence Characteristics
The dataset (without outgroups) comprises 139 sequences of 1143 nt length; 254 sites are variable and 197 parsimony-informative. Variable sites are documented in (Table 1) which clearly indicates the positions, which are informative and characteristic for each taxon.

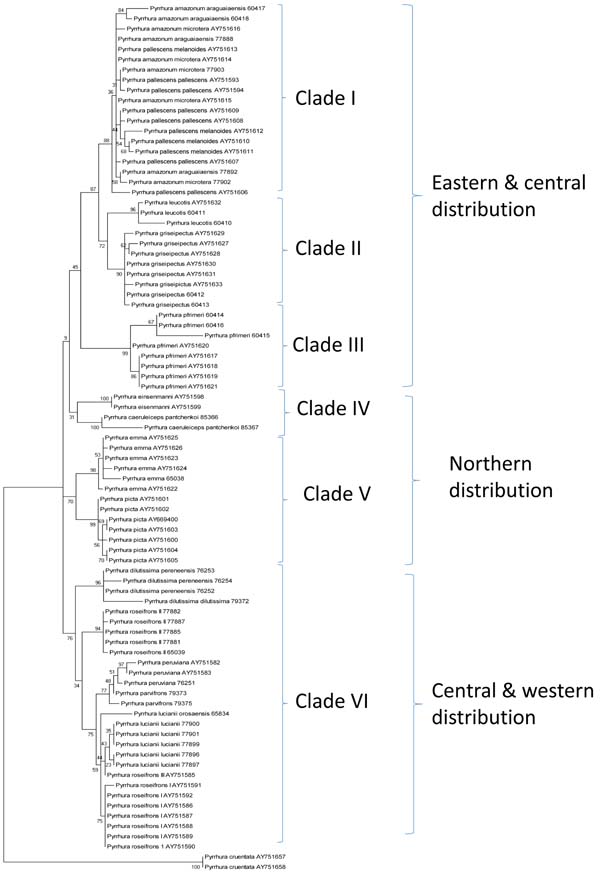 |
Fig. (3) Phylogenetic tree. Molecular Phylogenetic Analysis by Maximum Likelihood. The evolutionary history was inferred by using the Maximum Likelihood method based on the Tamura-Nei model [33Tamura K, Nei M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol Biol Evol 1993; 10(3): 512-26. [PMID: 8336541] ]. The tree with the highest log likelihood (-3141.3109) is shown. Robustness of nodes was assessed by bootstrap analysis (300 bootstrap replications). |
3.2. Phylogenetic Analyses
Phylogenetic trees were reconstructed with Maximum Likelihood (Fig. 3 ).
).
The following clades could be recovered with good bootstrap support: Clade I (comprising amazonum and pallescens), Clade II (with leucotis and griseipectus), Clade III (with 2 lineages of pfrimeri), Clade IV (with eisenmanni and caeruleiceps), Clade V (with emma and picta) and Clade VI (with peruviana, dilutissima, parvifrons, lucianii and 3 lineages of roseifrons). The positions of eisenmanni and caeruleiceps are unresolved on the basis of bootstrap values. However, most clades can be identified by specific and characteristic nucleotide substitutions Table 1, even if they lack high bootstrap support, because of a low number of nucleotide substitutions.
Clades I to III describe taxa with an eastern and central distribution in northern South America, clades IV and V show a northern and clade VI a western and central distribution pattern (Fig. 3).
3.3. Cluster Analysis
The cluster analysis of morphological and ecological characters almost entirely confirmed the results of the species rank method Fig. (2). The values for taxa achieving species status lay between 0.08 and 0.1. All values below this indicated a subspecies according to the species rank method. The only exception arose with cuchivera; this form clustered with pallescens because of its obvious similarities.
3.4. Amazonum / Pallescens Group
Specimens Examined
Pyrrhura amazonum
Altogether, there were 25 preserved specimens of amazonum (north of the Amazon) and 87 of microtera (south of the Amazon) available for the assessment of this form.
Joseph [5Joseph L. Geographical variation, taxonomy and distribution of some Amazonian Pyrrhura parakeets. Ornith Neotrop 2002; 13: 337-63.] pointed out that a few amazonum specimens have red feathers on the bend of wing, which could reflect hybridisation with picta. However, in fact only two specimens had isolated red feathers. Otherwise, they showed no tendency of any kind towards picta, so that the morphological division between the two forms was very clear. None of the picta representatives showed tendencies to amazonum in their colouring.
Representatives of amazonum as well as microtera varied considerably in breast colour (but not in breast feather markings), undermining the case for a division into two subspecies. Joseph [5Joseph L. Geographical variation, taxonomy and distribution of some Amazonian Pyrrhura parakeets. Ornith Neotrop 2002; 13: 337-63.] pointed out, however, that microtera is on average smaller, and our measurements confirm this (Appendix 4). It has to be taken into account that the Amazon provides a natural barrier, which prevents gene-exchange, and microtera is spread over three river systems (Rio Tapajós, Rio Xingu and Rio Tocantins/Rio Araguaia). The average wing length of amazonum is 117.9 mm (n = 23). The specimens of microtera highlighted a clear west-east division. The measurements of birds from Rio Tapajós are 113.8 mm (n = 43), from Rio Xingu 111.3 mm (n = 13) and from Rio Tocantins/Araguaia 109.2 mm (n = 23). A molecular genetic comparison of the populations north and south of the Amazon was unfortunately not possible, so for the moment we regard it as premature to lump microtera with amazonum.
Following the appearance in Brazilian and European live bird collections of a clearly different form of the Pyrrhura amazonum group, supposedly originating from the Rio Tocantins, we extended our specimen examination to the most important Brazilian museums. This revealed that the species occurred along the Tocantins as well as the Rio Araguaia, an evidence, which had been largely overlooked. Pacheco and Olmos [34Pacheco JF, Olmos F. As aves do Tocantins 1: Região Sudeste. Revista Brasileira de Ornitologia 2006; 14: 85-100.], for example, cite P. pfrimeri in their list of birds of Tocantins, but not amazonum. TA was able finally to link three specimens from Santana do Araguaia, Faz. Fartura, Pará (MZUSP 89888, 89889 and 89890) to the unknown form and confirm this by several visits to the area concerned. It became evident that the form, named above as araguaiaensis, lives in the seasonal forest typical of the Mato Grosso, which extends locally to the river system of the Araguaia.
Pyrrhura pallescens
Altogether 90 specimens of this taxon were available to us. Despite the relatively large range and a significant variation in the breast feather markings and colouring, all specimens could immediately be assigned to pallescens by their broad, pointed breast feather edging, which extends to the lower cheeks. This makes a confusion with other taxa unlikely. Birds from Jacareacanga on the Rio Tapajós, which were identified by Ribas et al. [6Ribas CC, Joseph L, Miyaki CR. Molecular systematics and patterns of diversification in Pyrrhura (Psittacidae) with special reference to the picta-leucotis complex. Auk 2006; 123: 660-80.
[http://dx.doi.org/10.1642/0004-8038(2006)123[660:MSAPOD]2.0.CO;2] ] as snethlageae (now pallescens), belong to microtera (the authors confused Jacareacanga, Rio Tapajós, with Jacareacanga, Rio Bonito, where pallescens actually occurs).
Arndt [7Arndt T. Anmerkungen zu einigen Pyrrhura-Formen mit der Beschreibung einer neuen Art und zweier neuer Unterarten. Papageien 2008; 21: 278-86.] separated the birds from Rio Teles Pires at the mouth of the Rio Cristalino near to Alta Floresta and from the Rio Peixote de Azuvedo as the subspecies lucida, as they were paler in breast feathering and generally smaller, with more blue to the forehead. However, Gaban-Lima and Raposo [9Gaban-Lima R, Raposo MA. The status of three little known names proposed by Miranda-Ribeiro (1926) and the synonymization of Pyrrhura / snethlageae Joseph & Bates, 2002 (Psittaciformes: Psittacidae: Arinae). Zootaxa 2016; 4200: 192-200.
[http://dx.doi.org/10.11646/zootaxa.4200.1.10] [PMID: 27988647] ] showed that P. pallescens melanoides is the valid senior name for P. snethlageae lucida. Careful further investigation of the specimen material revealed that the entire pallescens population can be split into two large groups: representatives from the upper catchment area of the Rio Madeira and the Rio Juruena (pallescens: little or no blue on the forehead, larger, but smaller bill [15.0 mm; n = 27]) and birds of the upper Rio Tapajós and Rio Xingu (melanoides: marked blue forehead, smaller, but larger bill [16.1 mm; n = 15]). The Cohen's d value for the difference in bill size was 1.27. The paler breast was marked in melanoides, but was also present as a trend in pallescens.
Biogeographic and Ethological Analysis
As mentioned above amazonum and pallescens cannot be separated by sequence data although an identification is possible. In this regard, our extended data do not differ from what Ribas et al. [6Ribas CC, Joseph L, Miyaki CR. Molecular systematics and patterns of diversification in Pyrrhura (Psittacidae) with special reference to the picta-leucotis complex. Auk 2006; 123: 660-80.
[http://dx.doi.org/10.1642/0004-8038(2006)123[660:MSAPOD]2.0.CO;2] ] already found out. However, if ecological, vocal and ethological differences are taken into account, it becomes obvious that these outweigh the genetic aspect. Although, both amazonum and pallescens inhabit primarily moist forest, they show differences in the preferred altitudes Table (2). In addition, there is the fact that microtera/araguaiaensis in northern Brazil meet pallescens in a semi-circle for 1,500 km (Appendix 1, Map 1) without any recorded hybridisation between the two species. This situation would be extremely unusual for parrot subspecies and there is no comparable situation within the family. The separation is especially clear in the outermost part of northeastern Mato Grosso where the taxa remain strictly on their own river system, but approach each other to within a few kilometres.

In addition to the morphological differences in breast markings and size there is a vocal difference. The species differ in the tone pitch they emit. We have for comparison drawn on four sound recordings with comparable flock calls (microtera: XC11420, XC119952; pallescens: XC37963, XC118834), where without doubt further detailed investigation would be helpful.
A further indication of the proximity of two species is revealed by the comparison with P. perlata and P. lepida, whose ranges are virtually identical with those of pallescens and amazonum and which are clearly genetically two species. Helbig et al. [16Helbig AJ, Knox AG, Parkin DT, Sangster G, Collinson M. Guidelines for assigning species rank. Ibis 2002; 144: 518-25.
[http://dx.doi.org/10.1046/j.1474-919X.2002.00091.x] ] put forward such a comparison with sympatric species for the assessment of allopatric taxa.
Species Rank Analysis
A comparison of pallescens with amazonum shows the following: the taxa occur parapatrically (3), pallescens is significantly larger [Cohen‘s d winglength = 2.00; tail length = 1.25] (2), with a brown forehead (1), brown breast with a strong V marking and very broad edging (3), different voice (1) and different habitat (not scored). The total score is 10.
The comparison suggests that species status for pallescens is justified.
3.5. Leucotis / Griseipectus Group
Phylogenetic Analysis
The results for these two taxa in the paper by Ribas et al. [6Ribas CC, Joseph L, Miyaki CR. Molecular systematics and patterns of diversification in Pyrrhura (Psittacidae) with special reference to the picta-leucotis complex. Auk 2006; 123: 660-80.
[http://dx.doi.org/10.1642/0004-8038(2006)123[660:MSAPOD]2.0.CO;2] ] are not clear. As only captive birds were available to these authors, there is the possibility of misidentification. In addition, griseipectus was once (as a subspecies of P. leucotis) just as correctly identified as Pyrrhura leucotis. We have therefore analysed our own samples with indisputable identifications as well as representatives of griseipectus (AY751633, AY751634) from GenBank (although AY751634 was listed as leucotis).

Specimens Examined
Altogether, there were 42 preserved specimens of Pyrrhura leucotis and 23 of P. griseipectus available for assessing these forms, which were all clearly identifiable and displayed clear differences. A detailed description of the diagnositic characters can be found in Olmos et al. [36Olmos F, Martuscelli P, Silva R, Silva E. Ecology and habitat of Pfrimer's Conure Pyrrhura pfrimeri, with a reappraisal of Brazilian Pyrrhura leucotis. Ornith Neotrop 1998; 8: 121-32., 37Olmos F, Silva WA, Albano C. Grey-breasted Conure Pyrrhura griseipectus, an overlooked endangered species. Cotinga 2005; 24: 77-83.], which especially highlight the differences in bill shape. P. griseipectus has a longer, broader and deeper bill, which gives the species the appearance of a larger head. Our measurements confirm this impression: the average bill length of leucotis is 14.5 mm (12.9-15.7 mm, n = 31) and of griseipectus 15.8 (14.5-16.6 mm, n = 16). Our wing and tail measurements (Appendix 4) show that griseipectus has slightly shorter wings, but a noticeably longer tail.
Biogeographic and Ethological Analysis
Pyrrhura leucotis and griseipectus are not only genetically separated, which is plausible because of geographical distance (ca. 1,500 km) between them, but also have ecological and ethological differences Table (3). Thus in addition to the clear differences in the habitat type used and altitude, there are differences in vocal pitch, which Olmos et al. [38Olmos F, Albano C, Campos A, Girão W, Minns J. Proposal (403) to South American Classification Committee: Elevate Pyrrhura griseipectus to species rank 2009. Retrieved from: http://www.museum.lsu.edu/~Remsen/ SACCprop403.html 2009.] already indicated. For comparison we used two sound recordings with comparable flock calls (leucotis: XC85379; griseipectus: XC16808).
Species Rank Analysis
A comparison of leucotis with griseipectus provided the following results: the taxa occur allopatrically (0), with major genetic differences (3), griseipectus has a clearly stronger bill [Cohen‘s d culmen length = 2.44] (2) and longer tail [Cohen‘s d tail length = 0.98] (not scored), but shorter wing [Cohen‘s d wing length = 1.24] (1), and differs in plumage in breast colour and breast edging (3) as well as vocally (1) and in habitat (1). The total score is 11.
The comparison shows that species status for griseipectus is justified.
3.6. Pfrimeri Group
Phylogenetic Analysis
Five sequences were available to us from the GenBank on Pyrrhura pfrimeri as well as 3 of our own sequences.
Specimen Examined
We examined 9 specimens of this taxon, which were all clearly identifiable and showed well-defined morphological differences to all other taxa of the picta-leucotis complex.
Biogeographic and Ethological Analysis
Pyrrhura pfrimeri is regarded today as a valid species, which is confirmed both by our genetic analysis and by the morphology and ecology of the taxon. Pfrimeri is the only form of the complex to live in caatinga habitat with deciduous or semi-deciduous dry forest growing on limestone outcrops or limestone-derived soils [36Olmos F, Martuscelli P, Silva R, Silva E. Ecology and habitat of Pfrimer's Conure Pyrrhura pfrimeri, with a reappraisal of Brazilian Pyrrhura leucotis. Ornith Neotrop 1998; 8: 121-32.].
Species Rank Analysis
A comparison of pfrimeri with all other taxa of the picta-leucotis complex (apart from emma and auricularis) shows the following: the taxon occurs allopatrically (0), the differences are genetically exceptional (4), pfrimeri differs morphologically through its blue forehead colour, which extends to the nape (2), blue breast colour with narrow edging (2) and red-brown ear-coverts (2) as well as habitat (1). The total score is 11.
The comparison confirms species status for P. pfrimeri.
3.7. Eisenmanni / Subandina / Caeruleiceps Group
Phylogenetic Analysis
Just 2 sequences of P. eisenmanni (AY751598, AY751599) were available to us from GenBank and 2 of P. caeruleiceps of our own.
Specimens examined
Altogether 2 preserved specimens of Pyrrhura eisenmanni, 6 of P. subandina, 10 of caeruleiceps and 2 of pantchenkoi were available for the assessment of these forms, which all clearly showed their differences in colour (Table 4) and size (Appendix 4).

The subspecies P. caeruleiceps pantchenkoi was described by Phelps [39Phelps WH Jr. Una nueva especie y dos nuevas subespecies de aves (Psittacidae, Furnariidae) de la Sierra de Perijá cerca de la divisoria Colombo-Venezolana. Bol Soc Venez Cienc Nat 1977; 33: 43-53.]. Joseph [4Joseph L. Beginning an end to 63 years of uncertainty: The Neotropical parakeets known as Pyrrhura picta and P. leucotis comprise more than two species. Proc Acad Nat Sci Philadelphia 2000; 150: 279-92.], examining one of the two preserved specimens of pantchenkoi, established that this form does not differ from caeruleiceps. In succeeding publications [5Joseph L. Geographical variation, taxonomy and distribution of some Amazonian Pyrrhura parakeets. Ornith Neotrop 2002; 13: 337-63., 40Arndt T. Neue Erkenntnisse zur Systematik der Rotschwanzsittiche. Papageien 2004; 17: 352-356, 389-94.] pantchenkoi was generally treated as a synonym of caeruleiceps. In fact, however, the holotype of pantchenkoi (AMNH 73168), which is clearly an immature with dull, brownish plumage, and the (probably adult) example in the Colección Ornitológica Phelps (COP 73169), both show a clear difference from caeruleiceps in the lores and eye area: these are coloured dark brown whilst nine adult caeruleiceps in USNM clearly show a red colour. The only other immature example (USNM 372618) has dark brown lores and eye area but lacks the dark blue cheek colour of both pantchenkoi specimens. Another difference relates to the breast feathers, which are more greyish with a paler edging in pantchenkoi. Observations and photos from the field confirm these differences (R. Strewe, pers. comm.). It is therefore not appropriate to treat pantchenkoi as a synonym of caeruleiceps, but for the time being it seems advisable to classify it as a subspecies of Pyrrhura caeruleiceps because of the colour differences.
Biogeographic and Ethological Analysis
Pyrrhura eisenmanni is currently recognised as a valid species, but this does not always apply to subandina and caeruleiceps, which, for example, are listed by the “South American Classification Committee” along with emma as subspecies of Pyrrhura picta. The Zoological Nomenclature Resource (www.zoonomen.net) follows this classification, but separates emma as a species. This does not make sense, as all the results to date show that the genetic relationships of the taxa within the picta-leucotis complex are also congruent to their geographical proximity. Distinctive morphological, ecological and ethological differences suggest that subandina is not a subspecies of P. picta, as emma and caeruleiceps occupy ranges between them. Although we do not expect this, at most it would be possible that subandina (and caeruleiceps) is a subspecies of P. emma, whose species status is genetically supported. But if it would be accepted that subandina is not an independent species, then its distribution and morphological similarity rather would suggest a close relationship to eisenmanni. In fact, all five taxa (picta, emma, caeruleiceps, subandina and eisenmanni) are geographically isolated from each other and, as would be expected, also show ecological and ethological differences (Table 5).

As mentioned before it is possible that caeruleiceps is merely a subspecies of P. emma. But our genetic analysis already confirmed species status for both taxa, which have long been isolated from each other; the shortest distance between caeruleiceps and emma is some 350 km.
Apart from some differences in the habitat types occupied (significant differences exist only between picta and the rest of the mentioned taxa (Table 5) and altitudes there are differences in the tonal pitch of the calls between emma and eisenmanni as well as between caeruleiceps and eisenmanni. We have drawn on four sound recordings with comparable flock calls (eisenmanni: XC107007; emma: XC202466; picta: XC65210; source: www.xeno-canto.org; caeruleiceps: Lynx_24395_Luis Eduardo Urueña, Source: www.ibc.lynxeds.com), although further detailed investigation would also be necessary.
Species Rank Analyses
The comparison between subandina and eisenmanni shows that both occur allopatrically (0), subandina is distinctly smaller (1), differs morphologically through the brown breast colour with narrow edging (3), the light blue tinge to the forehead and the blue lower cheeks (2) and the dull brown ear-coverts (2) as well as habitat (not scored). The total score is 8.
The comparison of subandina with picta, emma and caeruleiceps shows that all occur allopatrically (0), subandina is the taxon with the smallest size [caeruleiceps/subandina: Cohen‘s d wing length = 1.10; tail length = 1.46] (1), differs morphologically through the brown forehead with only a light blue tinge (2), the brown breast colour with narrow edging (2) and the green bend of wing (3) as well as habitat (not scored). The total score is 8.
A comparison of eisenmanni with picta, emma and caeruleiceps shows the following: the taxa occur allopatrically (0), the differences are genetically exceptional (4), eisenmanni differs morphologically through the brown forehead (3), the brown breast colour with narrow edging (2) and the green bend to the wing (3) as well as habitat (not scored). The total score is 12.
A comparison of caeruleiceps with emma shows the following: the taxa occur allopatrically (0), the differences are genetically exceptional (4), caeruleiceps differs morphologically through the breast colour with broad edging (3), the red colour to the forehead and lores (2), the dull brown ear-coverts and the brown colour of the nape (1, not evaluated) as well as habitat (scored), but not vocally (0). The total score is 9.
The comparison of caeruleiceps with picta shows the following: the taxa occur allopatrically (0), the differences are genetically exceptional (4), caeruleiceps is smaller with a relatively longer tail {Cohen‘s d winglength = 1.20; tail length = 0.18} (1) and differs in plumage through the breast colour with broad edging (3), the red colour to the forehead and lores (2) and the brown nape and cheek colour (not evaluated) as well as habitat (1) and vocally (1). The total score is 12.
The comparisons show that species status is not only justified for eisenmanni, but also for subandina and caeruleiceps.
3.8. Picta / Emma Group
Phylogenetic Analysis
Of this group 7 sequences of P. picta and 6 of P. emma were available to us from GenBank, which were supplemented by our own sequence for emma.
Specimens Examined
Altogether there were available for the assessment of these taxa 165 specimens of Pyrrhura picta, 38 of emma, 73 of auricularis, which were all unequivocally identifiable as picta or emma/auricularis and exhibited clear morphological differences in colour (Table 6) as well as in size (Appendix 4). Due to the only small differences between emma and auricularis it is not always possible to determine the subspecies without knowing the bird's origin.
Examining the range of variation in P. picta was not the intention of this study, but the seven specimens from the Río Cuchivero in COP stood out because of their pointed V-markings on the breast feathers, which strongly resembled that of pallescens, and their smaller size (Appendix 4); Phelps and Phelps [45Phelps WH, Phelps WH Jr. Eleven new subspecies of birds from Venezuela. Proc Biol Soc Wash 1949; 62: 109-24.] gave the name chuchivera for the population from which these specimens came. The validity of the taxon is worth future investigation.

Joseph [4Joseph L. Beginning an end to 63 years of uncertainty: The Neotropical parakeets known as Pyrrhura picta and P. leucotis comprise more than two species. Proc Acad Nat Sci Philadelphia 2000; 150: 279-92.], to whom 27 preserved specimens of auricularis and just 2 emma were available, established that auricularis is not diagnosable and should be synonymised with emma. Our research, however, demonstrated that both taxa have diagnosable differences. For Zimmer and Phelps [46Zimmer JT, Phelps WH. Four new subspecies of birds from Venezuela. Am Mus Novit 1949; 1395: 1-9.] auricularis possesses larger and clearer white auricular patches, a slightly darker and less yellowish green on the back and flanks, and slightly longer bill, wings and tail. Our measurements support this (Appendix 4), although the differences in green tone are not as great as stated in the original description. In addition we measured the width of the ear patch, which resulted in emma in an average of 7.5 mm (6.0-10.5 mm; n = 34) and in auricularis an average of 9.8 mm (6.9-14.0 mm; n = 44). The Cohen‘s d width of the ear patch for males = 3.11, for females = 2.75.
Biogeographic and Ethological Analysis
As expected, the taxa also exhibit ecological and ethological differences (Table 3). Apart from the clear differences in the habitat type and altitudes occupied there are the differences in tone pitch of the calls between emma and picta. We have used two sound recordings with comparable flock calls (emma: XC202466, picta: XC65210), although again further detailed investigation would be necessary.
Species Rank Analysis
A comparison of picta with emma shows the following: picta and emma occur allopatrically (0), emma is smaller (1), the genetic differences to picta are medium (2), emma differs morphologically through the breast colour with narrow edging (3), the very broad blue band to the nape (1) and the blue lower cheeks (1) as well as habitat (1) and vocally (1). The total score is 10.
The comparison indicates that species status is justified for emma.
3.9. Roseifrons / Parvifrons / Peruviana / Dilutissima / Lucianii Group
Phylogenetic Analysis
From this group we had available: roseifrons 8 GenBank and 5 of our own sequences; parvifrons 2 of our own; peruviana 2 GenBank and 1 of our own; dilutissima 4 of our own and lucianii 6 of our own.
This is without doubt the most complicated group within the complex. It is divided into five sub-groups, which, however, only partially form clearly defined units:
- dilutissima (including pereneensis, described above)
- roseifrons II (probably Rio Juruá*)
- peruviana and parvifrons
- lucianii (including orosaensis, described above) and roseifrons III (Río Madre de Dios)
- roseifrons I (Río Ucayali / Río Amazonas)
*samples from captivity in Brazil of uncertain origin
Specimens Examined
Altogether available for the assessment of these taxa were 82 specimens of Pyrrhura roseifrons, 26 of parvifrons, 18 of peruviana, 5 of dilutissima, 2 (+ 2 living birds) of pereneensis, 29 of lucianii and 13 of orosaensis. Unique to this group is that two taxa are difficult or impossible to determine without knowledge of origin (dilutissima and peruviana as well as parvifrons and orosaensis). P. lucianii and roseifrons are clearly determined; however, roseifrons splits into several sub-groups depending on the river system, which is also partly reflected in their size (Appendix 4). Plumage differences of the taxa are shown in (Table 7).

Roseifrons
The genetic division of roseifrons into three populations is not surprising as their distribution is determined by the big river systems (Ucayali- Amazonas {I}, Juruá {II} and Madre de Dios {III}). While our data show that both the Peruvian populations are closely related, the Brazilian population {II} appears to have diverged genetically, which could be due to its inhabiting varzea and adjacent habitat, although we could find no differences in the plumage colour between the three populations. The representatives from southern Peru and Bolivia {III} are significantly smaller (Appendix 4). Genetically this population is also nearer to lucianii, which belongs to the same river system as roseifrons from the Río Ucayali. A species rank analysis, however, shows clearly that separation of the three populations is impossible at the moment. The comparison of I with II achieves 6 points, I with III 2 points and of II and III maximum 6 points.
In addition, there is the situation that a genetic comparison for roseifrons II was based on available samples from captive birds of uncertain origin, while for roseifrons III only one sample was available to us.
Once we have more data we will examine the roseifrons complex more closely, in particular to determine whether a split into two species might be justified.
Parvifrons
This taxon was described by Arndt [7Arndt T. Anmerkungen zu einigen Pyrrhura-Formen mit der Beschreibung einer neuen Art und zweier neuer Unterarten. Papageien 2008; 21: 278-86.] as a species. The author still considered there exists a western (Shanusi, Yurimaguas and Sarayacu) and an eastern population (Santa Cecilia, Quebrada Vainilla and Río Orosa), although he pointed out that the eastern population differs slightly from Pyrrhura lucianii. He accepted that representatives from Tarapoto, which had a broad red forehead but identical breast markings like parvifrons specimens from Yurimaguas, are intermediate with roseifrons and both taxa possibly even occur sympatrically along the Río Cushabatay.
Several visits and fieldwork in the area of the western population as well as the eastern population, where samples could be collected, have surprisingly shown that the birds from Santa Cecilia, Quebrada Vainilla and Río Orosa belong genetically to P. lucianii (see lucianii below). Birds from Yurimaguas to Tarapoto form one group, although the proportion of birds with a broad red forehead, which somewhat resemble roseifrons, increases the further one travels in the direction of Tarapoto [26Arndt T, Gonzales Pinedo H. Die Rotschwanzsittiche in der Cordillera Escalera, Peru. Papageien 2013; 26: 204-7.]. A renewed examination of the specimen material from Tarapoto (USNM 108269, LSUMZ 116350, SMF 26052-4) confirms that it does not represent roseifrons, but belongs with parvifrons.
This also applies to a series of five examples (LSUMZ 161562, LSUMZ 161563, LSUMZ 161564, MJPL 7749, MJPL 808) from the Río Cushabatay (77 and 84 km WNW from Contamana), which Arndt [7Arndt T. Anmerkungen zu einigen Pyrrhura-Formen mit der Beschreibung einer neuen Art und zweier neuer Unterarten. Papageien 2008; 21: 278-86.] still considered as representatives of roseifrons and parvifrons occurring sympatrically. After further examination of the skins, we consider them all as parvifrons and no longer accept that roseifrons and parvifrons co-occur, which would be unique within the picta-leucotis complex, but we assume that the taxa are parapatric and meet in the area of the confluence of Río Cushabatay and Río Uyacali.
It is still noteworthy that parvifrons lives in a white river sand area, a habitat that clearly differs from the southwest Amazon moist forest occupied by roseifrons as well as that by peruviana and dilutissima [26Arndt T, Gonzales Pinedo H. Die Rotschwanzsittiche in der Cordillera Escalera, Peru. Papageien 2013; 26: 204-7., 47Frasier CL, Albert VA, Struwe L. Amazonian lowland, white sand areas as ancestral regions for South American biodiversity: Biogeographic and phylogenetic patterns in Potalia (Angiospermae: Gentianaceae). Org Divers Evol 2008; 8: 44-57.
[http://dx.doi.org/10.1016/j.ode.2006.11.003] ].
Peruviana
This taxon was described as a species by Hocking, Blake and Joseph [5Joseph L. Geographical variation, taxonomy and distribution of some Amazonian Pyrrhura parakeets. Ornith Neotrop 2002; 13: 337-63.] and according to our research is the sister group of parvifrons, but differs from it very clearly in morphology. Joseph [5Joseph L. Geographical variation, taxonomy and distribution of some Amazonian Pyrrhura parakeets. Ornith Neotrop 2002; 13: 337-63.] as well as Arndt [7Arndt T. Anmerkungen zu einigen Pyrrhura-Formen mit der Beschreibung einer neuen Art und zweier neuer Unterarten. Papageien 2008; 21: 278-86.] accepted that there is a northern and southern population, which our data do not support (see under dilutissima).
The main range of peruviana lies in the lower Río Marañón and the Río Santiago valley and their tributaries. However, there are also strays in the area of Chyavitas (NHM1889.1.30.151, RMNH 42 Cat 3, RMNH 43 Cat 2) and Chamicuros (NHM1869.5.25.107-8, NHM1890.6.1.89, ZMB 19399). A female from Chyavitas (= Chayahuitas, today Challavitas) shows the typical plumage colour of peruviana, but has few little red feathers to the base of the bill, which demonstrates the relationship and geographic proximity to parvifrons. At the same time the three Chyavitas specimens support the distribution of peruviana southwards along the Andes slopes, although Chyavitas lies on the most easterly tributary of the Marañón and with some certainty provides the border to the neighbouring parvifrons population. It is probable that both taxa meet in this area as the distance between Chyavitas and the Río Paranapu, which belongs to the river system of parvifrons, is just 5 km.
It is more difficult to interpret the four typical specimens of peruviana from Chamicuros. They were collected at approx. 175 m, which is not the typical habitat and altitude of peruviana. The species normally inhabits mountain regions at altitudes up to approx. 1,350 m along the rivers, and only comes into the valleys at certain times of the year for a short period for the fruit ripening of certain food plants. This is the main reason why a search for the species is often unsuccessful and why even the local people do not know the bird. Chamicuros lies too far from the nearest Andes area (ca. 100 km) for seasonal foraging to occur. An incorrect description of the locality where the specimens were collected is rather unlikely, as all were collected by Edward Bartlett, who was active for a while in Chamicuros. In fact, there is naturally the possibility that the distribution of peruviana is not limited to the area of the lower Andes, but stretches along the Río Marañón to the Río Huallaga. Specimen material from this scarcely known sector of the Río Marañón is not available.
Dilutissima
This taxon was described by Arndt [7Arndt T. Anmerkungen zu einigen Pyrrhura-Formen mit der Beschreibung einer neuen Art und zweier neuer Unterarten. Papageien 2008; 21: 278-86.] as a subspecies of P. peruviana based on the duller and washed-out edging to the breast. Joseph [5Joseph L. Geographical variation, taxonomy and distribution of some Amazonian Pyrrhura parakeets. Ornith Neotrop 2002; 13: 337-63.] pointed out that the northern population of peruviana noticeably differed morphologically from the southern, which our examples also showed. Dilutissima is significantly larger (Appendix 4), which is indicated above all by the bill measurements. The average bill length of peruviana is 15.3 mm (14.0-17.1 mm, n = 13) and of dilutissima 16.9 (15.0-18.5 mm, n = 9). Taking sexual dimorphism into account, the differences are even more apparent: the average bill length of peruviana males is 15.5 mm (14.0-17.1 mm, n = 7) and of dilutissima males 17.0 (16.5-17.4 mm, n = 2), while the measurements of peruviana females is 15.1 mm (14.2-15.8 mm, n = 6) and of dilutissima females 16.8 (15.0-18.5 mm, n = 7).
Additional fieldwork in the area of the upper Río Ené (May 2009, February and August 2014) and along the Río Perené (August 2013), where blood samples were also taken, showed surprisingly on the one hand that dilutissima differs genetically from all other taxa of the group and on the other that the Río Perené population differs so clearly from the neighbouring Río Ené population, owing to its somewhat narrower breast edging and a variable red marking on the front of the forehead, that a subspecific separation is justified (see also the dilutissima rank analysis).
Lucianii
Joseph [5Joseph L. Geographical variation, taxonomy and distribution of some Amazonian Pyrrhura parakeets. Ornith Neotrop 2002; 13: 337-63.] brought clarity to the circumscription of lucianii. This made possible an exact determination of museum specimens as well as the birds in the wild, which in turn led to new information about the distribution of this taxon. The surprising result is that the distribution of lucianii is not just along the Rio Purus but also along the Rio Madeira, where the species meets P. pallescens. Specimens of both taxa exist from Porto Velho (lucianii: MN8297, MN 8298, MN 8960; pallescens: AMNH148193, MZUSP 37.926-29). It had long been accepted that pallescens existed along the Rio Madeira, but in fact it has proven to occur only along a comparatively short section of that river.
Specimens from Amazonas / Rio Purus and the Rio Madeira show no differences in colour or size.
Joseph [5Joseph L. Geographical variation, taxonomy and distribution of some Amazonian Pyrrhura parakeets. Ornith Neotrop 2002; 13: 337-63.] accepted that P. parvifrons (he addressed them in his paper as “no 6 birds” as parvifrons was not named at this time) consisted of two populations (Shanusi, Yurimaguas and Sarayacu as well as the region of Santa Cecilia, Quebrada Vainilla and Río Orosa), which are connected to each other, and accordingly their occurrence is squeezed between the populations of P. roseifrons and P. peruviana. Arndt [7Arndt T. Anmerkungen zu einigen Pyrrhura-Formen mit der Beschreibung einer neuen Art und zweier neuer Unterarten. Papageien 2008; 21: 278-86.] likewise attributed both populations to parvifrons, but pointed out that there is no evidence for this connection and that it has rather more to do with two isolated populations. He accepted that the eastern population was genetically influenced by P. lucianii and originally spread westwards from the mouth of the Río Tefé along the Amazon. The contact with P. roseifrons led to its displacement and isolation today. This would explain why the specimens of this eastern population no longer differ greatly from P. lucianii. The extremely limited occurrence of this population was caused by the absence of larger tributaries of the Amazon in this area, which would have facilitated its spread southwards.
Our genetic research has in fact indicated that the populations of Santa Cecilia, Quebrada Vainilla and Río Orosa must be allocated to P. lucianii or roseifrons and not parvifrons. The morphological differences to roseifrons are, however, clear. Additionally, roseifrons comes into contact with this population on the Río Orosa without any sign of hybridization, which would be unusual for subspecies. The morphological similarities with lucianii are, however, striking (Appendix A1 ), group 6, photos A1 and 6). Seven of the 11 specimens available to us from Río Orosa have somewhat broader breast edges than lucianii, while the remaining four as well as the two specimens from Quebrada Vainilla do not differ from lucianii. All have the strong red marking to the front of the forehead, which is missing only on four birds and appears to support a separation as subspecies orosaensis, described above.
), group 6, photos A1 and 6). Seven of the 11 specimens available to us from Río Orosa have somewhat broader breast edges than lucianii, while the remaining four as well as the two specimens from Quebrada Vainilla do not differ from lucianii. All have the strong red marking to the front of the forehead, which is missing only on four birds and appears to support a separation as subspecies orosaensis, described above.
Biogeographic and Ethological Analysis
As expected the representatives of this group also exhibit ecological and ethological differences (Table 8).

Apart from the differences in the habitat types and altitudes occupied there are differences in the tone pitch of the calls of roseifrons, peruviana and dilutissima to lucianii. There are plainly no differences between the three roseifrons populations. We have taken six sound recordings with comparable flock calls (roseifrons I: XC 34386; roseifrons II: XC164646; roseifrons III: XC 2984; pereneensis: XC 104839; lucianii: XC 74133; Source: www.xeno-canto.org; peruviana: own data, TA) although further detailed research would be necessary.
Species Rank Analyses
Comparison of Pyrrhura roseifrons with all other taxa of the group indicates that all are separate species with pereneensis as a subspecies of P. dilutissima and orosaensis as a subspecies of P. lucianii. The details of the comparisons are listed below.
Parvifrons Species Rank Analyses
A comparison of parvifrons with roseifrons shows the following: the taxa occur parapatrically (3), the genetic differences are minor (1), parvifrons is smaller with somewhat shorter wings, but has a longer tail {Cohen‘s d wing length = 0.51; Cohen‘s d tail length = 1.10} (2), they differ morphologically through the breast colour and breast edging (1) as well as in head colour (2) and habitat (1). The total score is 10.
A comparison of parvifrons with peruviana shows the following: the taxa occur parapatrically (3), there are no significant genetic differences (0), parvifrons has a longer tail {Cohen‘s d wing length = 0.05; Cohen‘s d tail length = 1.17} (1), they differ morphologically through the breast colour and breast edging (1) as well as head colour (2) and habitat (1). The total score is 8.
A comparison of parvifrons with dilutissima shows the following: the taxa occur allopatrically (0), there are exceptional genetic differences (4), parvifrons is much smaller, but has a relatively longer tail {Cohen‘s d wing length = 1.16; Cohen‘s d tail length = 0.07} (2), they differ morphologically through the breast colour and breast edging (2) as well as the head colour (1) and habitat (1). The total score is 10.
A comparison of parvifrons with lucianii shows the following: the taxa occur allopatrically (0), there are clear genetic differences (3), parvifrons is smaller with somewhat shorter wings, but has a longer tail {Cohen‘s d wing length = 0.22; Cohen‘s d tail length = 0.67} (2), they differ morphologically through the breast colour and breast edging (1) as well as the head colour (1) and habitat (1). The total score is 8.
A comparison of parvifrons with orosaensis shows the following: the taxa occur allopatrically (0), there are clear genetic differences (2), parvifrons is smaller with somewhat shorter wings, but has a longer tail {Cohen‘s d wing length = 0.51; Cohen‘s d tail length = 0.20} (2), they differ morphologically through the breast colour and breast edging (1) and in habitat (1). The total score is 6.
These analyses show that parvifrons reaches species status in comparison with four of the five compared taxa and only fails with orosaensis. This was to expect as the morphological similarities of both taxa are significant and other data (e.g. comparison of sound calls) are not available yet. Nevertheless, the sum of our analyses indicates that species status for parvifrons is justified.
Peruviana Species Rank Analyses
A comparison with peruviana with roseifrons shows the following: the taxa occur parapatrically (3), the genetic differences are minor (1), peruviana is much smaller {Cohen‘s d wing length = 1.05; Cohen‘s d tail length = 1.47} (1), they differ morphologically through the breast colour and breast edging (2) as well as head colour (3) and only marginally in habitat (0). The total score is 11.
A comparison of peruviana with dilutissima shows the following: the taxa occur allopatrically (0), the genetic differences are major (3), peruviana is smaller {Cohen‘s d wing length = 1.24; Cohen‘s d tail length = 0.56} (1), they differ morphologically through the breast colour and breast edging (1) as well as the subspecies pereneesis in the head colour (2) and only marginally in habitat (0). The total score is 7.
A comparison of peruviana with lucianii shows the following: the taxa occur allopatrically (0), the genetic differences are major (2), they show scarcely any size difference {Cohen‘s d wing length = 0.31; Cohen‘s d tail length = 0.03} (0), they differ morphologically through the breast colour and edging (2) as well as the head colour (1), vocally (1) and in habitat (1). The total score is 7.
This comparison as well as that with parvifrons (see there) indicates that species status for peruviana is justified.
Dilutissima Species Rank Analyses
A comparison of dilutissima with roseifrons shows the following: the taxa occur parapatrically (3), the genetic differences are major (3), dilutissima is somewhat larger, but has a somewhat shorter tail {Cohen‘s d wing length = 0.69; Cohen‘s d tail length = 0.23} (1), they differ morphologically through the breast colour and breast edging (2) as well as head colour (3) and marginally in habitat (0). The total score is 12.
A comparison of dilutissima with lucianii shows the following: the taxa occur allopatrically (0), the genetic differences are exceptional (4), dilutissima is somewhat larger {Cohen‘s d wing length = 1.01; Cohen‘s d tail length = 0.43} (1), they differ morphologically through the breast colour and edging (2) as well as head colour (1), vocally (1) and in habitat (1). The total score is 10.
A comparison of dilutissima with pereneensis shows the following: the taxa probably share a mixed zone in the area where the Río Ené and Río Perené meet (1), no genetic differences are apparent (0), both taxa are the same size {Cohen‘s d has not been calculated on account of the small sample size} (0), they differ morphologically through the breast edging (2) as well as the head colour (2), but not in habitat (0). The total score is 5.
These comparisons as well as those with parvifrons and peruviana (see above) indicate that the species status for dilutissima is justified, but pereneensis can be separated as merely a subspecies.
Lucianii Species Rank Analyses
A comparison of lucianii with roseifrons shows the following: the taxa occur parapatrically (3), the genetic differences are minor (1), lucianii is somewhat smaller, but has a somewhat longer tail {Cohen‘s d wing length = 0.36; Cohen‘s d tail length = 0.34} (1), they differ morphologically through the breast colour and edging (2) as well as head colour (3), vocally (1) and clearly in habitat (1). The total score is 12.
A comparison of lucianii with orosaensis shows the following: the taxa occur allopatrically (0), there are no genetic differences (0), both taxa are the same size, but lucianii has a shorter tail {Cohen‘s d wing length = 0.00; Cohen‘s d tail length = 0.60} (1), they scarcely differ morphologically in the breast colour and edging (0), but clearly in the forehead colour (2) and scarcely in habitat (0). The total score is 3.
These comparisons as those with parvifrons, peruviana and dilutissima (see above) indicate that species status for lucianii and subspecies status for orosaensis are justified.
Surely, the Pyrrhura picta-leucotis complex is one of the most difficult in South America. But we have already came some good steps ahead with our knowledge and understanding of the complex. Nevertheless, some basic understanding are still lacking and our paper is certainly to be considered as work in progress. Thus the paraphyly and polyphyly in clade 1 (amazonum/pallescens) cannot be satisfactorily explained. Though McKay and Zink [49McKay BD, Zink RM. The causes of mitochondrial DNA gene tree paraphyly in birds. Mol Phylogenet Evol 2010; 54(2): 647-50.
[http://dx.doi.org/10.1016/j.ympev.2009.08.024] [PMID: 19716428] ] found a common cause of paraphyly to be incomplete lineage sorting due to recent speciation, they also point out that the mtDNA analyses, as used in our research, is not useful for delimiting species and other criteria, e.g. DNA markers with a higher resolution, must be employed.
In clade 6 there is still a need for research. This applies not only to the roseifrons populations, which might consist of two species, but also to the populations that are located along the Andes between the ranges of P. parvifrons and P. dilutissima. We expect even one or another surprising result from this area.
Currently, it seems, as in Peru two major groups (roseifrons I/peruviana/parvifrons and roseifrons II/dilutissima/lucianii) stand in competition and during this displacement process they already genetically influenced each other. It remains to be seen, if this hypothesis is true, how far this process has progressed and e.g. to which group the roseifrons III population belongs.
The new data on the distribution of taxa in Brazil indicate that the whole Amazon basin is inhabited by the Pyrrhura picta group (in contrast to previous assumptions). Map 1) indicates the areas for which data are still missing. So, it is not known which species are located in the area southwest of Manaus, the distribution limit of amazonum and pallescens needs to be determined more accurately, and for lucianii data are missing from the southwest of the assumed range.
3.10. Taxonomic Results
The research shows that the Pyrrhura picta-leucotis group consists of six cladistic/main groups, which can each be regarded as superspecies:
Clade I: amazonum/pallescens
Calde II: leucotis/griseipectus
Clade III: pfrimeri
Clade IV: eisenmanni/subandina*/caeruleiceps (* not sequenced)
Clade V: picta/emma
Clade VI: roseifrons/parvifrons/peruviana/dilutissima/lucianii
Within clades I and VI the relationships are not entirely clear and a species or subspecies designation is not possible solely according to phylogenetic, morphological or ecological evidence but requires a combination of these and in individual cases support by ethological aspects.
We postulate a division of the Pyrrhura picta-leucotis complex into the following species and subspecies shown in (Fig. 4 ):
):
Clade I: Amazonum / Pallescens
1. Pyrrhura amazonum amazonum Hellmayr 1906
Hellmayr’s Parakeet
(Pyrrhura picta amazonum Hellmayr, 1906, Bull. Brit. Orn. Cl. 19: 8)
Distribution: North of Amazonas between Monte Alegre and Oriximiná, Pará, Brazil (Appendix 1, Map 1).
Pyrrhura amazonum microtera Todd 1947
Lesser Hellmayr’s Parakeet
(Pyrrhura picta microtera Todd, 1947, Annals Carn. Mus. 30: 335)
Distribution: Eastern Amazon area south of Amazonas along the Rio Tapajós from Santarem to Jacareacanga in the west eastwards over the lower Rio Xingu to Paraguminas and Imperatriz in the east, then south along the Rio Tocantins to Palmas (Appendix 1, Map 1).
Pyrrhura amazonum araguaiaensis ssp. nov.
Rio Araguaia Parakeet
Type: MZUSP 89888, adult male, Mato Grosso, Santana do Araguaia, Faz. Fartura
ZooBank registration: 8DE4AD95-AB48-4936-90AA-9D47D85DB66E.
Diagnosis
This new form shares with Pyrrhura amazonum a narrow blue forehead and lower cheeks colouring, V-shaped broad edging to the throat and breast feathers, and a green bend of wing, but it differs from all other subspecies through the dark brown-grey breast colouring and the broad whitish to dull brown edging to individual feathers, on average much paler than the other Pyrrhura amazonum representatives; thus it appears generally to be more contrasting. Also the forehead has a more pronounced broad blue stripe, which stretches to the back of the eye area; the ear-coverts are paler dull brown; and the bird is smaller than the other amazonum subspecies.
A similar breast edging is found in P. picta, which has a blue forehead and forecrown and a red bend of wing. P. pallescens is larger and with significant narrow V-shapes of the throat and breast feathers resulting in very broad edgings. Additionally, it has no or less blue to the forehead. P. lucianii is much darker in the head colouring, has no or nearly no blue to the forehead and narrower edgings to the throat and breast feathers. Most closely araguaiaensis resembles P. peruviana and P. dilutissima from Peru, which however are much larger, with less contrasting edging to the throat and breast feathers, and less blue to the forehead. All other taxa differ clearly by either having red on the bend of wing or on the head.
Description of Type
Basic colour green (Parrot Green, 260); head dark brown (darker Vandyke Brown, 121); forehead blue (darker than Sky Blue 66); area beneath eye reddish brown; a few blue feathers on the cheeks; ear-coverts pale dull brown (Pale Horn Color, 92); lower cheeks, neck and breast feathers dark greyish brown (Dark Grayish Brown, 20), becoming slaty grey on the lower breast; neck and breast feathers broadly edged whitish (brighter than Cream Color, 54) or dull yellow; abdomen, lower back, rump and upper tail feathers reddish brown (between Burnt Sienna, 132, and Brick Red, 132A); primaries and secondaries blue (Turquoise Blue, 65).
Size of type, flattened wing: 113 mm; tail: 116 mm; upper bill length: 13.4 mm
Distribution
Presently only known from the area of the middle Rio Araguaia and its tributaries from Pau D'Arco, Tocantins, and the neighbouring areas of Pará to Confresa, Mato Grosso, Brazil (Appendix 1, Map 1).
2. Pyrrhura pallescens pallescens* Miranda-Ribeiro 1926
Madeira Parakeet
(Pyrrhura luciani pallescens Miranda-Ribeiro, 1926. Arquivos do Museu Nacional, 28: 11)
Distribution: from Porto Velho, Rondônia, and Periquito, Amazonas, southeast to Río Paucerna, Bolivia, and Pyrineus-Pires, Mato Grosso, Brazil (Appendix 1, Map 1).
Pyrrhura pallescens melanoides* Miranda-Ribeiro 1926
Cristalino Parakeet
(Pyrrhura luciani melanoides Miranda-Ribeiro, 1926. Arquivos do Museu Nacional, 28: 11.)
Distribution
From Novo Progresso and the upper Rio Iriri, Amazonas, southwards over the area of Rio Teles Pires as far as Vera and Canarana, Mato Grosso, Brazil (Appendix 1, Map 1).
Remarks
As can be seen from Fig. (3) and (Table 1), amazonum (and its subspecies) and pallescens are extremely closely related; there are no common nucleotide substitutions which would help to separate the taxa in Clade I.
* The recently used names Pyrrhura snethlageae snethlageae Joseph and Bates [5Joseph L. Geographical variation, taxonomy and distribution of some Amazonian Pyrrhura parakeets. Ornith Neotrop 2002; 13: 337-63.] and Pyrrhura snethlageae lucida Arndt 2008 are both junior synonyms as explained by Gaban-Lima and Raposo [9Gaban-Lima R, Raposo MA. The status of three little known names proposed by Miranda-Ribeiro (1926) and the synonymization of Pyrrhura / snethlageae Joseph & Bates, 2002 (Psittaciformes: Psittacidae: Arinae). Zootaxa 2016; 4200: 192-200.
[http://dx.doi.org/10.11646/zootaxa.4200.1.10] [PMID: 27988647] ].
Clade II: Leucotis / Griseipectus
3. Pyrrhura leucotis* (Kuhl 1820)
White-eared Parakeet
(Psittacus leucotis Kuhl, 1820, Nova Acta Acad. Caes. Leop. Carol. 10: 21)
Distribution
Southeast Brazil south of the Rio Jequitinhonha, Bahia, south to Espírito Santo and southeast Minas Gerais; in earlier times also São Paulo (Appendix 1, Map 2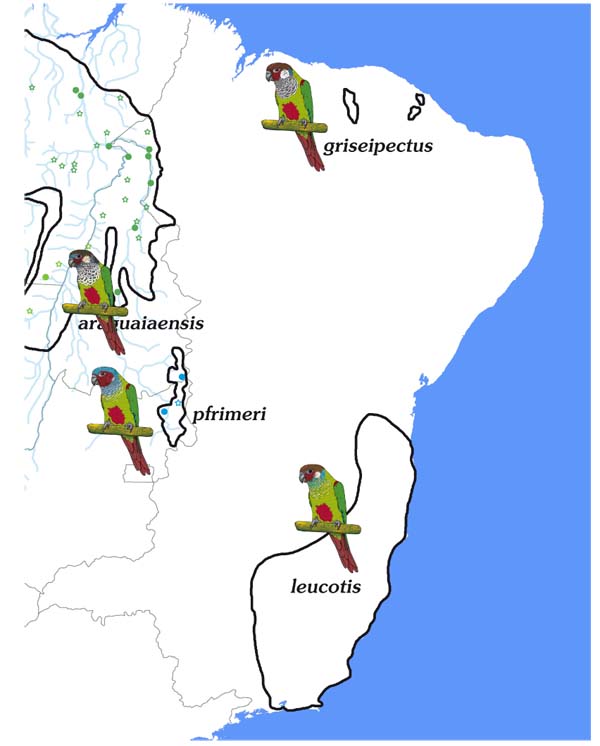 ).
).
 |
Fig. (4) Overview of the plumage patterns of all taxa of the Pyrrhura picta-leucotis complex. |
* Teixeira [50Teixeira DM. Revalidação de Pyrrhura anaca (Gmelin, 1788) do nordeste do Brasil (Psittaciformes: Psittacidae). Ararajuba 1991; 2: 103-4.] maintained that Kuhl’s P. leucotis is just a synonym for Pyrrhura anaca (Gmelin 1788) and therefore Gmelin’s name has precedence.
4. Pyrrhura griseipectus Salvadori 1900
Grey-breasted Parakeet
(Pyrrhura griseipectus Salvadori, 1900, Ibis: 672)
Distribution
Northeast Brazil in the Serra do Baturité and Quixadá, Ceará; formerly also in the Serra de Ibiapaba, Ceará, and the Serra Negra, Pernambuco (Appendix 1, Map 1).
Remarks
Leucotis and griseipectus cluster together in Clade II; they share a number of common nucleotide substitutions; they differ from each other by 9 nucleotide substitutions (Table 1).
Clade III: pfrimeri
5. Pyrrhura pfrimeri Ribeiro 1920
Maroon-faced Parakeet
(Pyrrhura pfrimeri Ribeiro, 1920, Rev. Mus. Paulista 12, pt. 2: 36)
Clade IV: eisenmanni / subandina* / caeruleiceps (* not sequenced)
6. Pyrrhura eisenmanni Delgado 1985
Azuero Parakeet
(Pyrrhura picta eisenmanni Delgado, 1985, Orn. Monogr. 36: 16)
Distribution
Western parts of the Azuero peninsula, Panama (Appendix 1, Map 3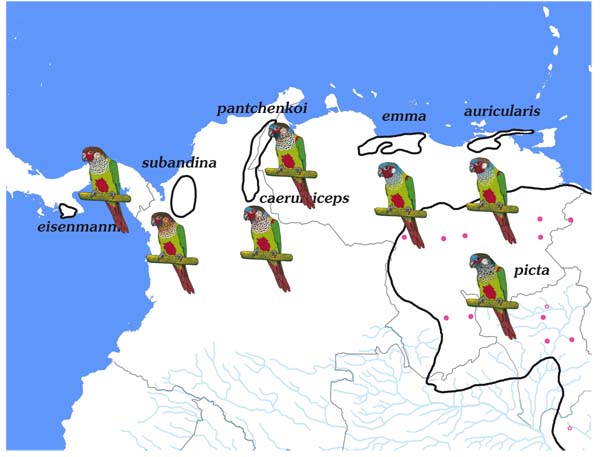 ).
).
7. Pyrrhura subandina Todd 1917
Sinú Parakeet
(Pyrrhura subandina Todd, 1917, Proc. Biol. Soc. Wash. 30: 6)
Distribution
Lower Sinú Valley, Sierra de Quimarí and Sierra de Murrucucú, northwest Colombia; probably already extinct (Appendix 1, Map 3).
8. Pyrrhura caeruleiceps caeruleiceps Todd 1947
Todd’s Parakeet
(Pyrrhura subandina caeruleiceps Todd, 1947, Ann. Carn. Mus. 30: 337)
Distribution
Spurs of the eastern Andes east of Aguachica, Cesar, and the adjoining Andes regions of Norte de Santander in northeast Colombia (Appendix 1, Map 3).
Pyrrhura caeruleiceps pantchenkoi Phelps 1977
Pantchenko’s Parakeet
(P. picta pantchenkoi Phelps, 1977, Bol. Soc. Venez. Cienc. Nat. 134: 45-47)
Distribution
Perijá mountains on the border of Colombia and Venezuela (Appendix 1, Map 3).
Clade V: Picta / Emma
9. Pyrrhura picta (P.L.S. Müller 1776)
Painted Parakeet
(Psittacus pictus P.L.S. Müller, 1776, Natursyst., Suppl., 1776: 75)
Distribution
Venezuela (Amazonas, Bolívar, in the Delta Amacuro south of the Orinoco), Guyana, Suriname, French Guiana, northern Brazil (Amapá, northeast Pará) (Appendix 1, Map 4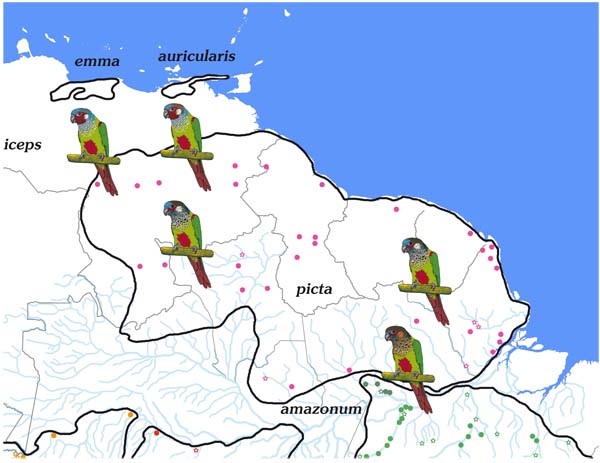 ).
).
10. Pyrrhura emma emma Salvadori 1891
Emma’s Parakeet:
(Pyrrhura emma Salvadori, 1891, Cat. Birds Brit. Mus. 20: 212 and 217, pl. 1)
Distribution
Coastal cordillera of northern Venezuela from Yacacuy and Carabobo to Miranda (Appendix 1, Map 4).
Pyrrhura emma auricularis Zimmer & Phelps 1949
Monagas Parakeet
(Pyrrhura emma auricularis Zimmer & Phelps, 1949, Am. Mus. Novit. no. 1395, p. 3.)
Distribution
Coastal cordillera of northern Venezuela from Anzoátegui to Sucre and northern Monagas (Appendix 1, Map 4).
Remarks
Emma and picta cluster together in Clade V; they share many nucleotype substitutions, but differ by 11 substitutions from each other, indicating that they constitute “good” species.
Clade VI: Roseifrons / Parvifrons / Peruviana / Dilutissima / Lucianii
11. Pyrrhura parvifrons Arndt 2008
Amazon Red-fronted Parakeet
(Pyrrhura parvifrons Arndt, 2008, Papageien 21: 279-280)
Distribution
Catchment area of Río Shanusi and Río Caynarachi from Yurimaguas, Loreto, to Tarapoto, San Martín, Peru; birds from the area of Sarayacu and the upper Río Cushabatay, both Loreto, belong with some certainty also to parvifrons (Appendix 1, Map 5).
12. Pyrrhura peruviana Hocking, Blake and Joseph 2002
Wavy-breasted Parakeet
(Pyrrhura peruviana Hocking, Blake & Joseph, 2002, Orn. Neotrop. 13: 356-357)
Distribution
Northwest Peru from Chiriaco along the Río Marañón and Río Santiago and their tributaries as far as Nantip in southeast Ecuador as well as the areas of Chyavitas and Chamicuros (Appendix 1, Map 5).
Remarks
According to sequence data, parvifrons and peruviana are almost identical and cannot be differentiated Fig. (3)
13. Pyrrhura lucianii lucianii (Deville 1851)
Deville’s Parakeet:
(Conurus lucianii Deville, 1851, Rev. et Mag. Zool [2Collar NJ. Psittacidae (Parrots). In: del Hoyo J, Elliott A, Sargatal J, Eds. Handbook of the birds of the world. Lynx Edicions, Barcelona 1997; pp. 280-477.], 3: 211)
Distribution
In the triangle of Alvaraes, Novo Aripuana, Brazil, and the Río Yata, Bolivia; possibly found along the Río Purus in the most easterly part of Peru (Appendix 1, Map 5).
Pyrrhura lucianii orosaensis ssp. nov.
Orosa Red-fronted Parakeet
Type: AMNH 230868, adult male; NE Peru, Orosa, Río Amazonas.
ZooBank registration: 8B35C59D-4125-4780-A3DA-7C68CA7DB2E5.
Diagnosis
Identifiable as conspecific with Pyrrhura lucianii by its dark head colouring caused by the nearly completely lack of blue to the forehead, slightly darker reddish brown cheeks and dull dirty brown to dark brown ear-coverts, with narrow throat and breast feather edging, but differs by a variable red forehead colouring, which is limited to fewer red feathers on the base of the bill or in a few (presumably young) birds is completely missing.
P. parvifrons is almost identically coloured, but the feather edging to the breast and throat is on average broader, the red part of the forehead much more extensive, and the reddish brown cheeks and the ear-coverts are paler. P. peruviana and P. dilutissima (including the new subspecies, described below which also has red to the forehead, but much more extensive) have broader edging to the throat and breast feathers and blue to the forehead. P. snethlageae is larger and with significant narrow V-shapes on the throat and breast feathers resulting in very broad edgings. P. amazonum has a broader edging to the throat and breast feathers and blue to the forehead. All other taxa differ clearly by either having red to the bend of wing or to the area around the eyes.
Description of Type
Basic colour green (Parrot Green, 260); small forehead patch red (Geranium, 12); bordering forehead area with faint blue tinge (darker than Sky Blue 66); head dark brown (darker Vandyke Brown, 121); upper cheeks dark reddish brown; a few blue feathers on the cheeks; ear-coverts dirty greyish-brown; lower cheeks, throat and breast feathers dark brown (Dark Grayish Brown, 20); throat and breast feather edging narrow and whitish (brighter than Cream Color, 54) or yellowish; abdomen, lower back, rump and upper tail feathers reddish brown (between Burnt Sienna, 132, and Brick Red, 132A); primaries and secondaries blue (Turquoise Blue, 65).
Measurements of Type
Flattened wing: 122 mm; tail: 119 mm; upper bill length: 15.7 mm
Distribution
Santa Cecilia region and Quebrada Vainilla along the Amazon to the mouth of the Río Orosa and along this river (Appendix 1, Map 5).
Remarks
Orosaensis differs from lucianii by 7 nucleotide substitutions Table (1); in the phylogram this taxon clusters basal to the lucianii/roseifrons clade. Its phylogenetic position is thus unresolved and needs to be further clarified.
14. Pyrrhura roseifrons (G.R. Gray, 1859)
Red-crowned Parakeet:
(Conurus roseifrons G.R. Gray, 1859, List Birds Brit. Mus., pt. 3, sect. 2: 42)
Distribution
São Paulo de Olivença and Rio Javarí, Brazil, southwest to eastern Peru; the southern population stretches from Itahuania, Peru, south to Teoponte, Bolivia (Appendix 1, Map 5).
Remarks
Sequence data indicate the existence of 3 lineages in this complex. Lineages I and III are closely related, whereas lineage II clusters basal to peruviana/parvifrons/lucianii/roseifrons I, III Fig. (3). Roseifrons II differs by 8 nucleotide substitutions from roseifrons I and III Table (1), suggesting that the latter two represent a new Pyrrhura species.
15. Pyrrhura dilutissima dilutissima Arndt 2008
Río Ené Parakeet
(Pyrrhura peruviana dilutissima Arndt, 2008, Papageien 21: 280)
Distribution
Area of Río Ené from the mouth of the Río Perené south to the area around Kimbiri and Luisiana in the cordillera Vilcabamba, central Peru (Appendix 1, Map 5).
Pyrrhura dilutissima pereneensis ssp. nov.
Río Perené Parakeet
Type: MJPL 26713; adult male; Sondobeni, Junín, central Peru; collector Peter Hocking.
ZooBank registration: 382A3388-2A37-481B-A695-B96F7AE34E31.
Diagnosis
Typical for Pyrrhura dilutissima is a combination of a broad, pale yellow edging of the breast feathers, the blue forehead colouring and a green bend of the wing. These features are also found in pereneensis. It differs from dilutissima by a variable red forehead colouring, which range from a few red feathers on the base of the bill (presumable in females and young birds) to nearly a complete red centre of the forehead and a slightly narrower edging to the throat and breast feathers.
P. parvifrons and orosaensis have also a red colouring to the forehead, but the brown head colouring is paler and the feather edging to the breast and throat is more clearly indicated. It appears thereby to be less dull and washed out than in pereneensis. P. peruviana has broader and less washed out edging to the throat and breast feathers and lacks the red to the forehead. P. pallescens has significant narrow V-drawings of the throat and breast feathers resulting in very broad edgings. P. amazonum lacks the red to the forehead, is smaller and has more blue to the forehead. All other taxa differ clearly by either having red to the bend of wing or to the area around the eyes.
This subspecies differs from all the other taxa descibed here by a combination of a broad, pale yellow edging of the breast feathers, the blue forehead colouring with a variable narrow red marking and a green bend to the wing. The feather edging is not so clearly indicated as in peruviana, and the edging to the breast thereby appears to be much duller and more washed out. This form differs from dilutissima by a somewhat narrower breast feather edging and the red marking to the forehead, which is lacking in young birds.
Description of the Type
Basic colour green (Parrot Green, 260); head dull brown (Vandyke Brown, 121); forehead light blue (darker than Sky Blue 66) with a narrow red patch on the base of the bill (Geranium, 12); cheeks under the eyes reddish brown; a few blue feathers on the cheeks; ear-coverts pale dull brownish (darker than Pale Horn Color, 92); lower cheeks, throat and upper breast feathers brown (Grayish Brown, 20), becoming green in the lower breast feathers; throat and breast feathers broadly edged pale cream or dull yellow; abdomen, lower back, rump and upper tail reddish brown (between Burnt Sienna, 132, and Brick Red, 132A); primaries and secondaries blue (Turquoise Blue, 65).
Measurements of Type
Flattened wing: 124 mm; tail: 122 mm; upper bill length: 17.4 mm
Distribution
Area of lower Río Perené and its tributaries, central Peru (Appendix 1, Map 5).
Remarks
This new form is genetically very close to the race dilutissima; because we had only a single specimen of d. dilutissima, it is difficult to say if the sequence differences are common with d. dilutissima or due to individual sequence variation.
APPENDIX 1. MAP AND ILLUSTRATION

 |
Map. (2) Distribution areas of Pyrrhura pfrimeri, P. griseipectus, P. leucotis and part of P. amazonum. |
 |
Map. (3) Distribution areas of Pyrrhura eisenmanni, P. subandina, P. caeruleiceps, P. emma and part of P. picta. |
 |
Map. (4) Distribution areas of Pyrrhura emma, P. picta and part of P. amazonum. |




APPENDIX 5. PHOTOS
All photos from T. Arndt unless stated otherwise

 |
Fig. (A5_2) Group 5: photo 1 - picta (captivity); photo 2 - emma (captivity). |

APPENDIX 6. SPECIMENS EXAMINED
P. amazonum amazonum (25). All localities in Brazil. Cachoeira do Tronco: UFRJ 3657; Lago Cuiteua (= Cuipeua): MZUSP 15739-42; Monte Alegre: MPEG 4412, MPEG 4862-3, MPEG 12998-9, SMF 26055-6, SMF 26058, ZMB no2, ZMB 311931, UFRJ 3653, UFRJ 3656; Obidos: AMNH 414612-4, MZUSP 10651-2, ZMB 311930, ZMB 311933; Río Trombetas: MZUSP 63957.
P. amazonum microtera (84). All localities in Brazil. Altamira, Río Xingu: MZUSP 85378-9; USNM 572507-8; Araguatins: MPEG 36722-23; MZUSP 52367-72, MZUSP 72262; Aramanai, Río Tapajós: MZUSP 32064; Aramathena, Río Tocantins: MPEG 5524; Boa Vista, Río Iriri: MPEG 10628; Brabo, Río Tapajós: AMNH 285834-9; Carajas: MPEG 36721; Caxricatuba, Río Tapajós: AMNH 285833, MZUSP 20395; Corrego das Panelas, Río Xingu: NM 44812; Curua-una: UFRJ 25748; Diamantina: USNM 121052-5; Estreito, Río Tocantins: MPEG 15584; Fazienda Sopezal, Tocantins: MZUSP 80358; Fordlandia: MZUSP 58204, MZUSP 62598, MZUSP 63032, MZUSP 63035, MZUSP 64819, MZUSP 64821, MZUSP 64828-9, MZUSP 66531; Goiatins, Río Tocantins: MZUSP 86371-7; Gorotire, Río Fresco: MZUSP 42673-5; Monte Christo, Río Tapajós: MZUSP 46408; Para: ZMB 30138; Parna, Río Tapajós: MPEG 4866; Santarem: AMNH 50216, AMNH 50222, MZUSP 3416-7, MZUSP 10645-8; Senador José PorfiRío, Río Xingu: MPEG 55586-9; Taperinha: MTD 30853-4; Tauari, Río Tapajós: AMNH 285823-32; Tucuruí, Río Tocantins: MPEG 36154; Urucurituba, Río Tapajós: MZUSP 58233, MZUSP 64820; Villa Braga, Río Tapajós: ZMB 6538, ZMB 311.932.
P. amazonum araguaiaensis (3). All localities in Brazil. Santana do Araguaia, Faz. Fartura: MZUSP 89888-90.
P. pallescens pallescens (67). All localities in Brazil unless stated otherwise. Allianca, Río Madeira, AMNH 474676; Arredores Forte Principe da Beira: UFRJ 37823; Cachoeira Nazaré: FMNH 330240, MPEG 39384-93, MPEG 39385; Calama: AMNH 474677-82, ZSM 576; Costa Marques: UFRJ 37788; Maruins, Río Machados: SMF 25934-6; ZSM 1000-2, ZSM 1037-38; Monicori: MPEG 57513; Periquo, Río Aripuana: MZUSP 62367-75; Porto Velho: AMNH 148193, MZUSP 37,926-9; Río Aripuana, Foz do Río Guariba: UFRJ 30920-33; Río Paucerna, Bolivia: LSUMZ 136840-1; Roosevelt River: AMNH 127347-9; São José do Río Claro: MZUSP 30, MZUSP 47; São Vicente, Río Machados: ZSM 1063.
P. pallescens melanoides (23). All localities in Brazil. Aldeira Kamaiura Bawá: UFRJ 30695; Alta Florest, Río Teles Pires, boca Río Cristalino: MPEG 51295-304; Cachimbo: MZUSP 38320-2; Fazienda São José, Río Peixote: MPEG 33505-6; Jacaré, Alto Xingu: UFRJ 30696; Paranato, Marg. E. Teles Pires: MZUSP 81753-4; Pyrineus-Pires: UFRJ 3651; Río Iriri: UFRJ 3654-5; Vera: MZUSP 75879.
P. leucotis (42). All localities in Brazil. Bahia: AMNH 474694, USNM 115223-5; Baixo Girardi: AMNH 317275; Casajeiras, Río Gongojuy: AMNH 241752; Colatina Municipio: MZUSP 24562; Esperito Santo: MZUSP 6406-8; Itabuna: MZUSP 10165; Lagoa Juparana: AMNH 317276-81; Linhares: USNM 368164-5; Río [de Janeiro]: AMNH 474693; Río Doce, Esperito Santo: AMNH 139941, MZUSP 6734; Río Doce, Minas Geraes: MZUSP 24808, MZUSP 24810-17; Río Itaunas: MZUSP 34495; Río Jucurucu: MZUSP 13988, MZUSP 13990; Río Muriae: MZUSP 27269-70; Río São José: MZUSP 28103-7; Serra Palhao, Bahia: MZUSP 13989.
P. griseipectus (23). All localities in Brazil. Ceara: ZMB 31/19935; Guaramiranda, Serra do Baturite: MZUSP 11221-2, MZUSP 11227; Serra do Baturite: MZUSP 41509-17, USNM 370348-9; captivity: MZUSP 78447, MZUSP 83520; Quixada: AMNH 241753, AMNH 241755-59.
P. pfrimeri (9). All localities in Brazil. Barra do Río São Domingo, Goias: MZUSP 15765, MZUSP 15767, MZUSP 15769; captivity: MZUSP 92581; Nova Roma, Río Parana: AMNH 474695-96; Pana Brava, Nova Roma, Goias: MZUSP 15766, MZUSP 15768, MZUSP 15770.
P. eisenmanni (2). Locality in Panama. Las Piraguales del Corteso de Tonosu, Los Santos Prov.: AMNH 824933, AMNH 824181.
P. caeruleiceps caeruleiceps (10). All localities in Colombia. Below Airoca: USNM 372610; Guamalito: USNM 372611-19.
P. caeruleiceps pantchenkoi (2). Locality in Venezuela. Frontera, Sierra de Perija: AMNH 73168, COP 73169.
P. subandina (6). Locality in Colombia. Nazaret, Río Sínu, 12 miles NW Tierra Alta: USNM 410662-7.
P. picta (163). All localities in Guyana unless stated otherwise: Abacy River: USNM 587505; Acari Mountains, North side: USNM 625107; Albina, Maroni River, Surinam: RMNH 213655-6; Amapa, Río Tracajatuba, 25 km E Ferreira Gomes, Brazil: USNM 51469093; Annai: AMNH 474668; Baramita: USNM 586307; Berbice: RMNH Cat. 7-8; British Guyana: RMNH 3713; Camp Jaime Benitez, Cerro El Negro, Venezuela: AMNH 42336, USNM 444105-6; Camp Rosenberg: MZUSP 6491, MZUSP 6496; Cayari Island, Uassa Swamp, Amapa, Brazil: AMNH 233718; Cano Demerara: AMNH 474669; Cavaimaken, Río Paragua, Bolívar, Venezuela: COP 81753; Cerro Parú, Falda Oeste, Caño Laguna, Amazonas, Venezuela: COP 47056-61; Cerro Yaví, Cerro, Falda Este, Amazonas, Venezuela: COP 37579, COP 37585; Cerro Yaví, Cerro, Falda Oriental, Amazonas, Venezuela: COP 37580-4; Clevelandia, Amapá, Brazil: UFRJ 3658; Commetewane, Surinam: RMNH 213651-2; El Dorado, Bolívar, Venezuela: COP 17445-6; El Palmar, Bolívar, Venezuela: COP 17686-7; El Venamo, Bolívar, Venezuela: COP 67836-44; Guiana: ZMB 66.609; Igarapé Novo, Amapá, Brazil: MPEG 16548, MPEG 28534, MPEG 29120, MPEG 29426; Ilha de Maiacá, Roraima, Brazil: MPEG 39014-6; Imataca, Sierra de, Altiplanicie de Nuria, Bolívar, Venezuela: COP 16838-41; Jaua, Meseta, Río Marajano, Bolívar, Venezuela; COP 65790; Kabalebo, Surinam: RMNH 37888; Kabel, Surinam: RMNH 14559; Kwata, Surinam: AMNH 474661-64; La Paragua, Bolívar, Venezuela: COP 15874-5, COP 15877, COP 15879-85; La Pricion, Caura R., Venezuela: AMNH 474656-59; La Union, Caura, Venezuela: AMNH 76096-105; Makalla: ZMB no 2-4; Manoa, Delta Amacuro, Venezuela: COP 1271-2; Marapanim, Brazil: RMNH 1152, RMNH 1158; Maura River, French Guiana: AMNH 233717; Mirico Creek, Surinam: RMNH 35209; Mt Duida, Foothills Camp, Venezuela: AMNH 272377; Mt Marahuacia, Venezuela: USNM 444105-06; Mucajai, Roraima, Brazil: MPEG 45655-63; Paramaribo, vicinity of, Surinam: AMNH 313405-06, AMNH 474667, ZMB 1857.2.8; Perú, El Callao, Bolívar, Venezuela: COP 74280; Pied Saut, French Guiana: AMNH 233719; Phedra, Surinam: RMNH 35779; Quonga: USNM 145687-92; Río Caura, Salto Pará, Bolívar, Venezuela: COP 25029-33; Río Chiguao, Cerro Tigre, Bolívar, Venezuela: COP 15876, COP 15878, COP 15886; Río Cuchivero, Cerro El Negro, Cumbre, Bolívar, Venezuela: COP 42330-4; Río Cuchivero, Cerro El Negro, Falda Este, Bolívar, Venezuela: COP 42335-6; Río Cunucunuma, Culebra, Amazonas, Venezuela: COP 74877; Río Maracá, Amapá, Brazil: MPEG 19880, MZUSP 42862-3; Río Paru de Leste, Para, Brazil: UMSP 31539; Río Surumu, Roraima, Brazil: MZUSP 73286; Río Tracajatuba, Amapá, Brazil: USNM 514690-4; Río Vatumá, Amapá, Brazil: MPEG 43645; Seco, Mte Duida, Venezuela: AMNH 270445; Sierra Pacaraima, Cerro Urutaní, Bolívar, Venezuela: COP 73507; Sierra Parima, Frontera 3, Amazonas, Venezuela: COP 71418-9; Suapure, Venezuela: AMNH 474660; Surinam: AMNH 474665-66; Valle de los Monos, Mte Duida, Venezuela: AMNH 272371, AMNH 272373-75; Venezuela: ZMB 10136; Vila Velhado Capicore, Amapá, Brazil: MPEG 19877-9; Villa Nova, Para, Brazil: MPEG 28151.
P. emma (38). All localities in Venezuela. Acevedo, Distrito, Cerro Negro, Falda Este, Miranda: COP 31255-8; Carenero, Miranda: COP 8413-4; El Guapo, Miranda: COP 80751; Hacienda Panchito, Yaracuy: COP 5312-6; Sierra de Aroa, Bucaral, Yaracuy: COP 26716; Sierra de Aroa, Cerro El Candelo, Yaracuy: COP 63892-6; Sierra de Aroa, Finca El Jaguar; Quebrada El Charal, Yaracuy: COP 77268-74; San Casimiro, Aragua: COP 80223; San Casimiro, Cerro Golfo Triste, Aragua: COP 19011-5; San José de Los Caracas, Distrito Federal: COP 18003-5; Los Caracas, DF: AMNH 387973; Sta Lucia, Miranda: USNM 389625.
P. emma auricularis (73). All localities in Venezuela. Bergantín, Quebrada Bonita, Anzoátegui: COP 15504-8; Caripe, Monagas: COP 79145-6; Caripe, Cerro Negro, Monagas: COP 22830-4; Caripe, Cuenca Media Río, Monagas: COP 78471; Cristobal Colon: AMNH 120353-61, COP 44368-73; Cueva del Gúacharo, Monagas: COP 78776; El Tigre, Anzoátegui: COP 82335; Falda Sur, Cerro Humo: AMNH 41008; Irapa, Cerro Humo, Falda Oeste, Sucre: COP 44140; Irapa, Cerro Humo, Falda Sur, Sucre: COP 56811-4, COP 41006-11; Irapa, Cerro Humo, Falda Norte, Sucre: COP 56815-6; Los Altos, Sucre: COP 15166-8, COP 85659; Macuro, Puerto Cristobal Colón, Sucre: COP 40630-9; Quebrada Seca, South of Cumana: AMNH 474683-90; Río Neveri: AMNH 188162-65; Trinidad: USNM 145684-85; Uquire, Sucre: COP 75806.
P. roseifrons (82). All localities in Peru unless stated otherwise. Abra Aguachini, ca 30 km SW Puerto Bermudez: LSUMZ 130085-7; Aguas Calientes, Contamana: FMNH 320233-4, MJPL 7561, MJPL 7578, MJPL 7584; Aguas Calientes, 75 km E Shintuya: FMNH 397723; Carabaya, Yahuarmayo: ZSM 1910; Cerro de Pantiacolla, above Río Palatoa: FMNH 320430; Cerro Tahauyo, SE slope, ca 65 km ENE Pucallpa: LSUMZ 156182-3, MJPL 4422; Conchapen: FMNH 285078; Hacienda Santa Elena, ca 35 km NE Tingo Maria: LSUMZ 72175; Itahuania: FMNH 222875-80; Joao Pessoa, Río Juruá, Brazil: FMNH 183717; MZUSP 16260-1, MZUSP 20385, MZUSP 20393-4, MZUSP 20476, MZUSP 21045, MZUSP 22483; Juanjui, 86 km SE of: LSUMZ 170686-8; La Pampa, Sandia: AMNH 145927; Lima, Alto Río Moa, Brazil: MPEG 52706; Nevati: FMNH 297882-3; Nusiniscato: FMNH 208169-70; Palcazu: AMNH 474703-4, MJPL 17037, MJPL 17046; Pucallpa: MJPL 1799, MZUSP 21048, MZUSP 64912; Requena Ucayali: AMNH 40678-9; Río Ene at mouth of Río Shesha, west bank, ca 65 km ENE Río Pucallpa: LSUMZ 156184-6, MJPL 11443-4, MJPL 11625; Río Javari, Amazonas, Brazil: MPEG 16965-77, UFRJ 29054; Río Pachitea: MJPL 19170; Porongaba, Río Juruá, Brazil: MPEG 48046; Río Juruá, Brazil: AMNH 474705, MZUSP 3502-4, SMF 26057; Río Shesha, casaRío Abujao: MJPL 4702; São Paulo de Olivença, Río Solimões, Brazil: CM 32984; Vila Taumatuigo, Río Juruá, Brazil: MPEG 28152; Yurinaqui Alto: FMNH 278311, MJPL7573, MJPL 30587.
P. parvifrons (26). All localities in Peru. Contamana, ca 77 km WNW of, Río Cushabatay: LSUMZ 161563-4; Sarayacu, R. Ucayali: AMNH 237723; Río Cushabatay, north-east bank, 84 km WNW of Contamana: LSUMZ 161562, MJPL 808, MJPL 7749; Shanusi: AMNH 474697-702, MPEG 28153, MZUSP 2275, SMF 26050-1, SMF 88363/2193, USNM 145680-1, ZMB 20267-8; Tarapoto: SMF 26052-4, USNM 108269; Tarapoto road to Yurimaguas, km 20: LSUMZ 116350.
P. peruviana (18). All localities in Peru. Bashium, Río Kagka: LSUMZ 33998; Betel, Río Santiago: FMNH 296580; Chamicuros: NHM1869.5.25.107-8, NHM1890.6.1.89, ZMB 19399; Chyavitas: NHM1889.1.30.151, RMNH 42 cat 3, RMNH 43 cat 2; Cordillera EL Condor: MJPL 11857; Puerto Galilea, Río Santiago: FMNH 278312-4; Oberer Amazonas: SMF 26059; Río Santiago: AMNH 406881; Tutinum, Río Cenepa: LSUMZ 33996-7; Villa Gonzalo, Río Santiago: MJPL 6765.
P. dilutissima dilutissima (5). All localities in Peru. Luisiana, 2 km E, Cordillera Vilcabamba: AMNH 819816; Luisiana, 6 km E, Cordillera Vilcabamba: AMNH 819871; Pumorini, Kimbiri Alto: MJPL (2 skins without number); Río Ene at mouth of R. Quipachiari: AMNH 820834.
P. dilutissima pereneensis (2). All localities in Peru. Sondobeni, Junin: MJPL 21081, MJPL 26713.
P. lucianii lucianii (29). All localities in Brazil unless stated otherwise. Abuna, Río Madeira: MZUSP (2 skins without number); Abuna, Barreiro 11: MZUSP (3 skins without number); Arimã, Río Purús: CM 93531, CM 93581; Brasil: RMNH 335 Cat 1; Caicara, Barreiro 19: MZUSP (skin without number); Humaita: AMNH 474675; Hyutanahan, Río Purús: CM 86406, CM 86546-50, CM 86620-1, CM 86660, CM 86964; Mujica Nova: MZUSP 76578-80; Oberer Amazonas: MTD 3486; Porto Velho, Río Madeira: UFRJ 8297-8, UFRJ 8960; Río Yata, Bolivia: ZMB 431.276; Santa Isidoro, Tefé: AMNH 308975.
P. lucianii orosaensis (13). All localities in Peru. Orosa River: AMNH 230866-74, MJPL 114613-4; Quebrada Vainilla, east bank, ca 10 km SSW mouth Río Napo: LSUMZ 114613-4.
ZOOBANK REGISTRATION
ZooBank registration number for the article: B518CB93-D834-4C73-96E5-87CF414765BE.
CONFLICT OF INTEREST
The authors confirm that this article contents have no conflict of interest.
ACKNOWLEDGEMENTS
We would like to thank the curators and staff of the following collections, who allowed TA to research existing material and supported us in our work: American Museum of Natural History, New York (P. Sweet, P. Hart), Field Museum of Natural History, Chicago (J. Bates), United States National Museum, Washington (J. Dean), Louisiana State University Museum of Natural Science, Baton Rouge (J. V. Remsen, S. Cardiff, D. L. Dittmann), Carnegie Museum of Natural History, Pittsburgh (S. Rogers), British Museum of Natural History, Tring (R. Prys-Jones), Museu de Zoologia da Universidade de São Paulo, São Paulo (L. F. Silveira), Museu Nacional, Río de Janeiro (M. A. Raposo), Museu Goeldi, Belém (A. Aleixo), Museum Alexander Humboldt, Berlin (S. Frahnert), Forschungsinstitut und Museum Senckenberg, Frankfurt am Main (G. Mayr), Staatliches Museum für Tierkunde in Dresden, Dresden (M. Päckert), Zoologische Staatssammlung München (M. Unsöld), Museo de Historia Natural “Javier Prado” de la UNMSM, Lima (I. Franke) und Colección Ornitológica Phelps, Caracas (M. Lentino, M. Martínez).
Special thanks to L.F. Silveira (Museu de Zoologia da Universidade de São Paulo, São Paulo) as well as W. Kiessling, M. Reinschmidt and D. Waugh (Fundación Loro Parque) for their generous provision of test material. We thank Hedi Sauer-Gürth (IPMB) who carried out the DNA isolation, amplification and sequencing.
We also acknowledge Servicio Nacional Forestal y de Fauna Silvestre (SERFOR) for permission to collect blood samples.
We would like to thank the following for their active assistance, helpful information and valuable comments: T. Pittman, N. Collar, N. Bahr, H. Schnitker, P. Hocking, A. Fergenbauer-Kimmel, R. Niemann, R. Strewe, M. Braun, C. Manderscheid, M. Schaefer, R. Prinz, J. Asmus, R. Wüst, P. Wolf, P. Teles, H. Gonzales Pinedo, G. Mayr, K.-H. Lambert, C. Navarro and M. Sanchez.
REFERENCES
| [1] | Forshaw JM, Cooper WT. Parrots of the world. 3rd (rev) ed. London: Blandford Press 1989. |
| [2] | Collar NJ. Psittacidae (Parrots). In: del Hoyo J, Elliott A, Sargatal J, Eds. Handbook of the birds of the world. Lynx Edicions, Barcelona 1997; pp. 280-477. |
| [3] | Juniper T, Parr M. Parrots: A guide to the parrots of the world. Robertsbridge, UK: Pica Press 1998. |
| [4] | Joseph L. Beginning an end to 63 years of uncertainty: The Neotropical parakeets known as Pyrrhura picta and P. leucotis comprise more than two species. Proc Acad Nat Sci Philadelphia 2000; 150: 279-92. |
| [5] | Joseph L. Geographical variation, taxonomy and distribution of some Amazonian Pyrrhura parakeets. Ornith Neotrop 2002; 13: 337-63. |
| [6] | Ribas CC, Joseph L, Miyaki CR. Molecular systematics and patterns of diversification in Pyrrhura (Psittacidae) with special reference to the picta-leucotis complex. Auk 2006; 123: 660-80. [http://dx.doi.org/10.1642/0004-8038(2006)123[660:MSAPOD]2.0.CO;2] |
| [7] | Arndt T. Anmerkungen zu einigen Pyrrhura-Formen mit der Beschreibung einer neuen Art und zweier neuer Unterarten. Papageien 2008; 21: 278-86. |
| [8] | Del Hoyo J, Collar NJ. HBW and BirdLife International Illustrated checklist of the birds of the World. Lynx Edicions, Barcelona: Non-Passerines 2014; Vol. 1. |
| [9] | Gaban-Lima R, Raposo MA. The status of three little known names proposed by Miranda-Ribeiro (1926) and the synonymization of Pyrrhura / snethlageae Joseph & Bates, 2002 (Psittaciformes: Psittacidae: Arinae). Zootaxa 2016; 4200: 192-200. [http://dx.doi.org/10.11646/zootaxa.4200.1.10] [PMID: 27988647] |
| [10] | Rheindt FE, Székely T, Edwards SV, et al. Conflict between genetic and phenotypic differentiation: the evolutionary history of a lost and rediscovered shorebird. PLoS One 2011; 6(11): e26995. [http://dx.doi.org/10.1371/journal.pone.0026995] [PMID: 22096515] |
| [11] | Filardi CE, Smith CE. Molecular phylogenetics of monarch flycatchers (genus Monarcha) with emphasis on Solomon Island endemics. Mol Phylogenet Evol 2005; 37(3): 776-88. [http://dx.doi.org/10.1016/j.ympev.2005.02.007] [PMID: 16291094] |
| [12] | Mayr E. Systematics and the origin of species. Cambridge, Mass: Harvard Univ. Press 1942. |
| [13] | Gill FB. Species taxonomy of birds: Which null hypothesis? Auk 2014; 131: 150-61. [http://dx.doi.org/10.1642/AUK-13-206.1] |
| [14] | Johnson NK, Remsen JV Jr, Cicer C. Resolution of the debate over species concepts in ornithology: A new comprehensive biologic species concept. Proc Int Ornithol Congr. 1470-82. |
| [15] | Tobias JA, Seddon N, Spottiswoode CN, Pilgrim JD, Fishpool LD, Collar NJ. Quantitative criteria for species delimitation. Ibis 2010; 152: 724-46. [http://dx.doi.org/10.1111/j.1474-919X.2010.01051.x] |
| [16] | Helbig AJ, Knox AG, Parkin DT, Sangster G, Collinson M. Guidelines for assigning species rank. Ibis 2002; 144: 518-25. [http://dx.doi.org/10.1046/j.1474-919X.2002.00091.x] |
| [17] | Brown WL, Wilson EO. Character displacement. Syst Zool 1956; 5: 49-64. [http://dx.doi.org/10.2307/2411924] |
| [18] | Cracraft J. Speciation and its ontology: The empirical consequences of alternative species concepts for understanding patterns and processes of differentiation. In: Otte D, Endler JA, Eds. Speciation and its consequences. Sunderland, MA: Sinauer Associates 1989; pp. 28-59. |
| [19] | Zink RM, McKitrick MC. The debate over species concepts and its implications for ornithology. Auk 1995; 112: 701-19. |
| [20] | Winker K. Subspecies represent geographically partitioned variation, a goldmine of evolutionary biology, and a challenge for conservation. Ornithol Monogr 2010; 67: 6-23. [http://dx.doi.org/10.1525/om.2010.67.1.6] |
| [21] | Bush MB, Oliveira PE. The rise and fall of the refugial hypothesis of amazonian speciation: A paleoecological perspective. Biota Neotrop 2006; 6(1) [http://dx.doi.org/10.1590/S1676-06032006000100002] |
| [22] | Haffer J, Prance GT. Climatic forcing of evolution in Amazonia during the Cenozoic: On the refuge theory of biotic differentiation. Amazoniana 2001; 16: 579-608. |
| [23] | Prance GT. A Review of the phytogeographic evidences for Pleistocene climate changes in the Neotropics. Ann Mo Bot Gard 1982; 69: 594-624. [http://dx.doi.org/10.2307/2399085] |
| [24] | Haffer J. Avian speciation in tropical South America. Cambridge: Nuttall Ornithological Club, Harvard University 1974. |
| [25] | Klauke N, Schaefer HM, Bauer M, Segelbacher G. The influence of natural habitat structure and habitat disturbance on the fine-scale genetic structure of an endemic Neotropical parrot. Mol Ecol submitted |
| [26] | Arndt T, Gonzales Pinedo H. Die Rotschwanzsittiche in der Cordillera Escalera, Peru. Papageien 2013; 26: 204-7. |
| [27] | Girão W, Campos A, Albano C. Das Schutzprojekt für den Salvadori-Weißohrsittich. Papageien 2008; 21: 29-32. |
| [28] | Smithe FB. Naturalist’s color guide. New York: American Museum of Natural History 1975. |
| [29] | Alström P, Ranft R. The use of sounds in avian systematics and the importance of bird sound archives. Bull Bull Brit Orn Cl 2003; 123: 114-35. |
| [30] | Wink M, El-Sayed A-A, Sauer-Gürth H, Gonzalez J. Molecular phylogeny of owls (Strigiformes) inferred from DNA sequences of the mitochondrial cytochrome b and the nuclear RAG-1 gene. Ardea 2009; 97: 209-19. [http://dx.doi.org/10.5253/078.097.0425] |
| [31] | Wink M, Sauer-Gürth H. Phylogenetic relationships in diurnal raptors based on nucleotide sequences of mitochondrial and nuclear marker genes. In: Chancelor RD, Meyburg B-U, Eds. Raptors Worldwide. Berlin: WWGBP 2004; p. 483498. |
| [32] | Kumar S, Stecher G, Tamura K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol Biol Evol 2015. submitted [PMID: 27004904] |
| [33] | Tamura K, Nei M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol Biol Evol 1993; 10(3): 512-26. [PMID: 8336541] |
| [34] | Pacheco JF, Olmos F. As aves do Tocantins 1: Região Sudeste. Revista Brasileira de Ornitologia 2006; 14: 85-100. |
| [35] | Fund W. Ecoregions of Brazil. Retrieved from: http://www.eoearth.org/view/article/177836 2012. a |
| [36] | Olmos F, Martuscelli P, Silva R, Silva E. Ecology and habitat of Pfrimer's Conure Pyrrhura pfrimeri, with a reappraisal of Brazilian Pyrrhura leucotis. Ornith Neotrop 1998; 8: 121-32. |
| [37] | Olmos F, Silva WA, Albano C. Grey-breasted Conure Pyrrhura griseipectus, an overlooked endangered species. Cotinga 2005; 24: 77-83. |
| [38] | Olmos F, Albano C, Campos A, Girão W, Minns J. Proposal (403) to South American Classification Committee: Elevate Pyrrhura griseipectus to species rank 2009. Retrieved from: http://www.museum.lsu.edu/~Remsen/ SACCprop403.html 2009. |
| [39] | Phelps WH Jr. Una nueva especie y dos nuevas subespecies de aves (Psittacidae, Furnariidae) de la Sierra de Perijá cerca de la divisoria Colombo-Venezolana. Bol Soc Venez Cienc Nat 1977; 33: 43-53. |
| [40] | Arndt T. Neue Erkenntnisse zur Systematik der Rotschwanzsittiche. Papageien 2004; 17: 352-356, 389-94. |
| [41] | Fund W. Ecoregions of Colombia 2012b. Retrieved from: http://www.eoearth.org/view/article/ 51cbf3e67896bb431f6ae6a8 2012. b |
| [42] | Botero-Delgadillo E, Paez CA. Estado actual del conocimiento y conservación de los loros amenazados de Colombia. Conservación Colombiana 2011; 14: 86-151. |
| [43] | Forshaw JM. Parrots of the world: An identification guide. Princeton, NJ and Oxford, UK: Princeton University Press 2006. |
| [44] | Hilty SL. Birds of Venezuela. Princeton: Princeton University Press 2003. |
| [45] | Phelps WH, Phelps WH Jr. Eleven new subspecies of birds from Venezuela. Proc Biol Soc Wash 1949; 62: 109-24. |
| [46] | Zimmer JT, Phelps WH. Four new subspecies of birds from Venezuela. Am Mus Novit 1949; 1395: 1-9. |
| [47] | Frasier CL, Albert VA, Struwe L. Amazonian lowland, white sand areas as ancestral regions for South American biodiversity: Biogeographic and phylogenetic patterns in Potalia (Angiospermae: Gentianaceae). Org Divers Evol 2008; 8: 44-57. [http://dx.doi.org/10.1016/j.ode.2006.11.003] |
| [48] | Saundry P. Peru 2012. Retrieved from: http://www.eoearth.org/ view/article/ 51cbee9e7896bb431f699219 |
| [49] | McKay BD, Zink RM. The causes of mitochondrial DNA gene tree paraphyly in birds. Mol Phylogenet Evol 2010; 54(2): 647-50. [http://dx.doi.org/10.1016/j.ympev.2009.08.024] [PMID: 19716428] |
| [50] | Teixeira DM. Revalidação de Pyrrhura anaca (Gmelin, 1788) do nordeste do Brasil (Psittaciformes: Psittacidae). Ararajuba 1991; 2: 103-4. |
Endorsements
Browse Contents
Table of Contents
- INTRODUCTION
- MATERIALS AND METHODS
- RESULTS AND DISCUSSION